Effects of dietary tributyrin on intestinal mucosa development, mitochondrial function and AMTK-mTOR pathway in weaned pigs
Background
Piglets are often subjected to nutritional, physiological and immunological stresses during the weaning process [1]. It has been determined that weaning can quickly lead to the intestinal-mucosal injury, such as villus atrophy and crypt hyperplasia and further impair intestinal absorption function [2–5]. Butyrate, as a SCFA (short chain fatty acid), has gained much attention due to its rewarding effects on cellular energy metabolism and intestinal homeostasis [6]. Butyrate is rapidly absorbed across the luminal membranes of intestinal epithelial cells under the action of butyrate transporters [7]. Substantial evidence has reported that butyrate plays a potential role in affecting epithelial cell growth and differentiation, and repairing intestinal injury [8–10]. Tributyrin, containing three molecules of butyrate, has also been reported to maintain intestinal mucosa normal function [11, 12]. Therefore, it is necessary to study the beneficial effects of butyrate or tributyrin on intestinal development and absorption function in weaned piglets.
The intestine requires a lot of energy to renew and repair the damaged intestinal mucosa [13], and mitochondria are responsible for the main source of cellular energy [14]. Mitochondria are the primary site of reactive oxygen species (ROS) generation, and at the same time, they are highly susceptible to ROS [15]. Our previous study has found that weaning caused oxidative stress and severely impaired mitochondrial function [16]. Recently, Xing et al. [17] reported that in HepG2 cells, sodium butyrate alleviated oxidative injury and enhanced mitochondrial function. Besides, several papers reported that butyrate was effective in relieving intestinal oxidative stress [18, 19]. Nevertheless, no information is available about the effects of butyrate or tributyrin on oxidative stress and mitochondrial function of intestine in weaned pigs.
AMPK (AMP-activated protein kinase), a sensor of energy state, is responsible for regulating mitochondrial function and cell energy metabolism [20]. Activation of AMPK results in the inhibition of mTOR (mammalian target of rapamycin), a downstream regulator of AMPK that senses energy and stress [21]. Substantial evidence has suggested that the AMPKmTOR pathway acted as an important role in modulating oxidative stress [22–24]. However, the effect of butyrate or tributyrin on AMPK-mTOR pathway has not been reported so far.
Accordingly, we hypothesized that dietary tributyrin could enhance intestinal mucosa development, improve mitochondrial function and influence AMPK-mTOR signaling pathway of weaned pigs. Our objective was aimed at exploring the effect of dietary tributyrin on intestinal mucosa growth, mitochondrial function as well as AMPK-mTOR signaling pathway of weaned pigs.
 Methods
Methods
Experimental design
Seventy-two pigs (Duroc × Landrace × Yorkshire, body weight (BW) of 6.8 kg, weaned at 24±1 days of age), were divided into two treatments according to their initial BW and sex. Each treatment has six replicates of six pigs (3 barrows and 3 gilts). Two treatments received basal diet or the same diet supplemented with 750mg/kg tributyrin (provided by Shanghai Menon Feed Co. Ltd., Shanghai, China). The formula for the diets is in compliance with NRC (2012) (Table 1). Pigs had free to drink and feed.
Sample collection
After 14days, 6 pigs (3 barrows and 3 gilts) from each treatment (1 pig per pen) were euthanized with an ear intravenous injection of sodium pentobarbital (200mg/kg BW) (Sigma-Aldrich, St. Louis, MO) and the gastrointestinal tract was rapidly removed. Proximal jejunal samples were fixed in 4% paraformaldehyde for measurement of intestinal mucosa architecture. Segments of the proximal jejunum were immediately gained to isolate intestinal mitochondria. The mucosa from the adjacent jejunum were harvested and placed in liquid nitrogen for further analysis.
Measurement of intestinal mucosa architecture
Segments for morphological study were fixed in 4% paraformaldehyde and then embedded in paraffin wax. Sections of 5μm were cut and stained with hematoxylin and eosin. Crypt depth and villus height were measured in three intestinal cross sections with at least 10 welloriented crypt villus units for each cross-section using an image processing and analysis system (Leica Imaging Systems, Cambridge, UK) and averaged for each sample.
Measurement of mucosal DNA, RNA and protein
The jejunal DNA, RNA and protein were collected from snap-frozen mucosal samples, with TRI Reagent-RNA/DNA/Protein isolation reagent (TaKaRa Biotechnology, Dalian, China). The concentrations were measured according to Jiao et al. [25].
Determination of antioxidative enzyme activities
Antioxidant parameters, including superoxide dismutase (SOD), catalase (CAT), malondialdehyde (MDA) and hydrogen peroxide (H 2 O 2 ) in jejunum were determined with commercially available kits (Nanjing Jiancheng Bioengineering Institute, Nanjing, China).
Isolation of mitochondria
Fresh jejunal mucosa was prepared to isolate mitochondria. The process was in compliance with the rules of mitochondria isolation kit (Beyotime Institute of Biotechnology). After intestinal mucosa was homogenized in MSH buffer (10mmol/L HEPES, pH 7.5, containing 200 mmol/L mannitol, 70mmol/L sucrose, 1.0mmol/L egtazic acid and 2.0mg/mL serum albumin), the homogenate was centrifuged at 1000×g for 10 min at 4 °C. And then the collected supernatant was centrifuged at 3500×g for 10min at 4 °C to acquire mitochondrial pellet [26].
Mitochondrial reactive oxygen species (ROS) content
The fluorescent probes (2′,7′-dichlorohydro-fluorescein diacetate, DCFH-DA) were used for detecting mitochondria ROS content as previously described [27]. DCFHDA can pass through the cell membrane and be hydrolyzed into DCFH. ROS can oxidize non-fluorescent DCFH into DCF, which has a maximum absorption peak at an excitation wavelength of 485nm and an emission wavelength of 525 nm, and the intensity is proportional to the level of ROS. The fluorescence of DCF was assayed using a fluorescence microplate reader (FLx800, Bio-Tek Instruments, Inc.). The results were indicated as fold changes of the control.
Mitochondrial membrane potential (ΔΨm)
The changes of ΔΨm were assayed by ΔΨm assay kit with JC-1 (Beyotime Institute of Biotechnology). The isolated mitochondria were strained with JC-1 at 37°C for 15min, and then the fluorescence intensity was assayed using a fluorescence microplate reader (FLx800, Bio-Tek Instruments, Inc.). JC-1 monomer form (green) fluorescence was excited at 485nm, and the emission was detected at 530nm. JC-1 aggregate form (red) fluorescence was excited at 485nm, and emission fluorescence was detected at 590nm. The ratio of red and green fluorescence values was used to reflect the ΔΨm [16].
Determination of mitochondrial DNA (mtDNA) content
The mtDNA numbers relative to genomic DNA were assayed with co-amplification of the mitochondrial DNA-loop (mt D-loop) and the nuclear-encoded β-actin gene using RT- PCR according to the previous steps [28]. Total DNA was extracted from proximal jejunal mucosa using a TIANamp Stool DNA Kit (Tiangen Biotech, Beijing, China). For the mt D-loop the primers were forward 5′-GATCGT ACATAGCACATATCATGTC-3′, reverse 5′-GGTCCT GAAGTAAGAACCAGATG-3′. For β-actin the primers were forward 5′-CCCCTCCTCTCTTGCCTCTC-3′, reverse 5′- AAAAGTCCTAGGAAAATGGCAGAAG-3.
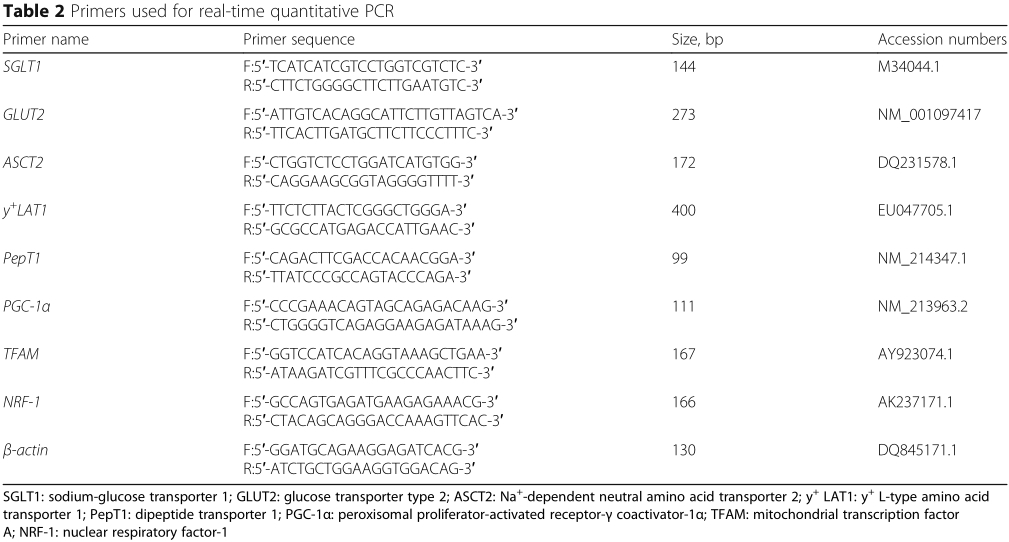
mRNA expressions of nutrient transporter genes and mitochondrial function-related genes
Total RNA of jejunal mucosa was obtained with the Trizol reagent (TaKaTa, Dalian, China) according to the kit’s direction. RNA concentration and purity were assayed by a Nano Drop spectrophotometer (ND-2000; NanoDrop Technologies, Wilmington, DE). Reverse transcription was performed with the PrimeScripte RT reagent kit (TaKaRa Biotechnology, Dalian, China). Quantitative RT-PCR was performed on a StepOne Plus real-time PCR system (Applied Biosystems, Foster City, CA) using SYBR Green Master mix (Promega, Madison, WI) as described by Liu et al. [29]. The genes include sodium glucose transport protein-1 (SGLT1), glucose transporter-2 (GLUT2), Na + -dependent neutral amino acid transporter 2 (ASCT2), y + L-type amino acid transporter 1 (y + LAT1), dipeptide transporter 1 (PepT1), peroxisomal proliferator-activated receptor-γ coactivator-1α (PGC-1α), mitochondrial transcription factor A (TFAM) and nuclear respiratory factor-1(NRF-1). Primers are exhibited in Table 2. The β-actin was used as a reference gene to normalize mRNA levels of each target gene. The 2 −ΔΔCt method was used to analyze the relative expression, calculated relative to the values from the control group.
Western blot assay
The steps are in accordance with the procedures of Hu et al. [5]. Sample in the jejunal mucosa was performed with SDS-PAGE and transferred to PVDF membranes. The membrane was blocked for 120 min at 25 °C, and then incubated with primary antibodies overnight at 4 °C. The membrane was rinsed and then incubated with the secondary antibodies for 60 min at 25 °C. The antibodies, including p-AMPK, AMPK, p-mTOR, mTOR, GAPDH and HRP-conjugated anti-rabbit Ab were purchased in Santa Cruz Technology Inc. (Santa Cruz, CA). The signal was displayed with Chemi Scope 3400 (ClinxScience Instruments, Shanghai, China) and the protein band value was counted with Image J analysis software.
Statistical analysis
Data were analyzed by Student's t-test with SPSS 22.0 statistical package (SPSS Inc., Chicago, IL). The significance value and a trend toward difference were set at levels of P< 0.05 and P <0.10, respectively.
 Results
Results
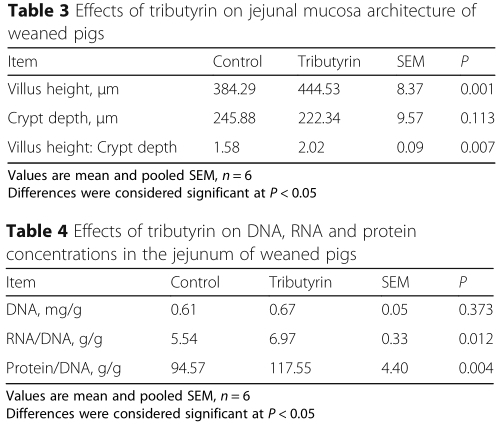
Effect of tributyrin on jejunal mucosa architecture of weaned pigs
Intestinal mucosa architecture is exhibited in Table 3.The weaning pigs fed with tributyrin showed higher (P < 0.05) jejunal villus height and villus height: crypt depth in comparison to the control. The crypt depth had no difference in two treatments (P >0.05).
Effect of tributyrin on jejunal DNA, RNA and protein of weaned pigs
Data about the concentrations of DNA, RNA and protein of weaned pigs are summarized in Table 4. Tributyrin increased (P <0.05) jejunal RNA/DNA ratio as well as protein/DNA ratio in weaned pigs. No alteration was observed in the DNA concentrations between the two groups.
Effect of tributyrin on jejunal mRNA expression of nutrient transporter of weaned pigs
As shown in Table 5, dietary tributyrin induced an increase (P <0.05) in SGLT1 and GLUT2 mRNA level, and had a trend (P <0.1) to raise mRNA level of ASCT2 inthe jejunal mucosa of weaned pigs.
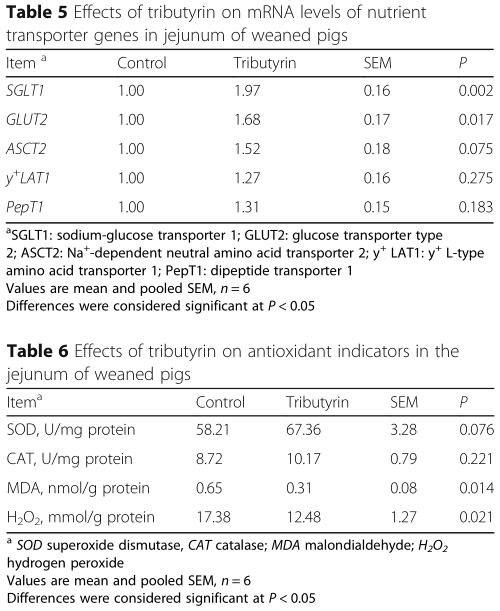 Effects of tributyrin on jejunal antioxidant indicators of weaned pigs
Effects of tributyrin on jejunal antioxidant indicators of weaned pigs
Table 6 indicates that the contents of MDA and H2 O2 in jejunum were reduced (P<0.05) in pigs fed tributyrin compared to those in control. However, no significant alteration (P >0.05) was observed in activities of SOD and CAT of jejunum between the two groups.
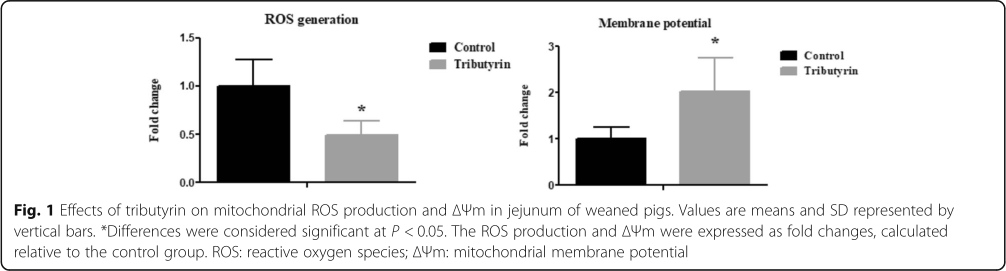
Effect of tributyrin on jejunal mitochondrial ROS and △Ψ of weaned pigs
Figure 1 shows that in comparison with the control, tributyrin significantly reduced (P <0.05) mitochondrial ROS level and raised (P <0.05) △Ψ.
Effect of tributyrin on jejunal mRNA expression of mitochondrial function-related genes of weaned pigs
Figure 2 shows that tributyrin increased (P<0.05) mtDNA copy numbers as well as mRNA levels of mitochondrial function-related genes, including PGC-1α, TFAM and NRF-1.
Effect of tributyrin on AMPK-mTOR signaling pathway in the jejunum of weaned piglets
Figure 3 shows the effect of tributyrin on the AMPKmTOR signaling pathway. Compared to the control, supplemental tributyrin effectively elevated (P<0.05) the ratio of the phosphorylated AMPK to total AMPK (p-AMPK/AMPK), while decreased (P<0.05) the ratio of the phosphorylated mTOR to total mTOR (p-mTOR/mTOR).
Discussion
Weaning pigs are often confronted with changes of intestinal architecture, including villus atrophy and crypt hyperplasia [5]. Butyrate, a volatile fatty acid, plays a significant part in enhancing intestinal mucosa architecture and promoting intestinal growth [30]. This current study demonstrated that tributyrin increased villus height as well as villus height: crypt depth, suggesting that weaning-induced injury to jejunal mucosa architecture was alleviated by addition with tributyrin. Similar to our results, Dong et al. [11] showed that in the intrauterine growth restriction pigs, dietary tributyrin increased villus height: crypt depth of the duodenum and jejunum, and lowed crypt depth of duodenum. The protein level, the ratio of RNA to DNA and protein to DNA in the intestinal mucosa are valuable biological parameters to assess intestinal growth and development [25, 31]. DNA concentration represents the mitosis rate to renew columnar epithelial cell, RNA/DNA and protein/DNA signifies cell efficiency and protein synthesis efficiency, respectively [25]. In this research, dietary tributyrin elevated the jejunal RNA/DNA and protein/DNA, which is beneficial for the small intestinal growth of weaned pigs. Similarly, another study found that SCFAs were trophic to the intestinal mucosa as evidenced by the increased DNA, RNA and protein concentration in intestinal mucosa of rats [32]. Monocarboxylate transporter 1 (MCT-1) plays an important role in the transport of monocarboxylates across cell membrane [33]. It has been identified in the small intestine [34] and colon [35]. An earlier research reported that in cultured colonic epithelial cells, treatment with sodium butyrate caused a concentration- and time-dependent upregulation of MCT1 mRNA and protein [36]. These results indicated that dietary tributyrin may enhance the expression of butyrate transporters in the jejunum, thereby promoting the intestinal mucosa development.
Weaned piglets need a huge amount of nutrients to meet the rapid renewal and growth of the intestine. The absorption of nutrients mainly depends on specific transporters that transport nutrients across the intestinal epithelium [4]. However, few researches studied the action of tributyrin in intestine absorption function of weaned pigs. Therefore, we observed the effects of tributyrin on mRNA expressions of intestinal nutrient transporters (SGLT1, GLUT2, ASCT2, y + LAT1, PepT1). SGLT1 is a rate-limiting step to transport glucose into the absorptive enterocyte, and GLUT2 transported glucose from intestinal cell into the blood circulation [37]. ASCT2 and LAT1, in charge for transporting neutral amino acid, have a principal part in the growth and proliferation of intestinal cell [38]. PepT1, widely existed in intestinal epithelial cell, is responsible to transport dipeptide from the lumen into enterocyte [39]. In present study, we found that the jejunal SGLT1 and GLUT2 mRNA abundances were elevated by tributyrin. The result was analogous with an in vitro study using Caco-2, which found that butyrate treatment elevated the mRNA level of GLUT2 [40]. Weaning piglets were confronted with innutrition and intestinal disorders due to the immature digestion and absorption systems [1], which might be part of the reason why tributyrin can promote the mRNA expressions of the nutrient transporters of weaning piglets. Accordingly, it is likely that tributyrin enhanced absorption function, thus transporting more nutrition to improve the intestinal mucosa architecture.
Several studies reported that tributyrin reduced intestinal oxidative stress in the ethanol-challenged colitis [41] and colon of dextran sodium sulphate-challenged mice [42]. However, there was little evidence about the antioxidant effect of tributyrin in pigs. Therefore, it is necessary to assess the effect of tributyrin on redox state in intestine of weaned pigs. Generally, CAT and SOD are regarded as two main antioxidant enzymes and have essential roles in the prevention of oxidative injury. Surprisingly, we found that dietary tributyrin had no significant effects on enzyme activities of CAT and SOD. However, Leonel et al. [42] found that tributyrin supplementation lowered the H 2 O 2 level, increased the SOD and CAT activities in dextran sodium sulphate-induced colitis of mice. Ma et al. [18] reported that after treatment with sodium butyrate, the levels of SOD, GSH-Px (glutathione peroxidase) and GSH (glutathione) increased while MDA level decreased in scratched IPEC-J2 cell. This discrepancy could be explained by the diverse trial conditions, animal species and animal model. MDA concentration in tissues and serum has been considered as biomarkers of oxidative injury [43]. In our study, the results showed that dietary tributyrin decreased MDA and H 2 O 2 level, which indicates that tributyrin relieved the intestinal oxidative stress of weaned pigs.

Weaning often causes oxidative stress and then damages mitochondria [16]. As a result, damaged mitochondria release more ROS and then lead to ΔΨm collapse by the formation of permeability transition [44]. Therefore, higher mitochondrial ROS and lower ΔΨm are considered to be the markers of mitochondrial dysfunction [45, 46]. Although prior studies have found that butyrate can reduce intestinal oxidative stress, no evidence established the effect on intestinal mitochondrial function of pigs. In this experiment, we observed that tributyrin reduced mitochondrial ROS level and increased ΔΨm of jejunum in comparison to the control. The findings suggested that tributyrin mitigated oxidative damage and enhanced mitochondria function. A previous study also found that, in high fat diet-induced obese mice, butyrate improved mitochondrial function reflected by the enhanced mitochondrial oxidative phosphorylation [47]. Li et al. [48] found butyrate inhibited the decrease of ΔΨm in LPS-challenged cow mammary epithelial cell. Davis et al. [49] reported that butyrate reduced the mitochondrial ROS by increasing proton leak through upregulation of uncoupling protein 2. Russo et al. [19] demonstrated that in intestinal epithelial cells, butyrate was effective in controlling the increase of ROS levels in response to lipopolysaccharide.
The mtDNA was known to function significantly in oxidative phosphorylation and normal mitochondrial function [50]. The mtDNA content can reflect the mitochondrial function [50]. In this research, the mtDNA content of piglets fed tributyrin was higher than pigs in control, which was supported by Xing et al. [17], who reported that sodium butyrate restored the H 2 O 2 -induced decrease of mtDNA copy number in HepG2 cells. The result indicates that dietary tributyrin is quite helpful in attenuating mtDNA damage and improving mitochondrial function of weaned piglets. The alteration of mtDNA content is in accordance with the changes of multiple transcriptional key regulators involved in mitochondrial biogenesis [51]. PGC-1α is proven to be a major regulator of mitochondrial biogenesis [52]. NRF-1 and TFAM are crucial for governing mtDNA replication and transcription during mitochondria biogenesis [53]. In the present study, the upregulated NRF-1, PGC-1α and TFAM mRNA abundances were observed in the jejunum of pigs fed tributyrin. Similarly, a previous study found that in oxidative injured HepG2 cells, sodium butyrate increased PGC-1α and TFAM mRNA levels [17]. Thus, tributyrin plays a relevant role in improving mitochondrial function of weaned pigs.
AMPK, a key energy sensor, functions in the regulation of cellular energy [54]. It has been shown that the activation of AMPK significantly modulates oxidative stress and mitochondrial function [55, 56]. Furthermore, AMPK activation inhibits mTOR, a serine/threonine protein kinase [21]. A large body of evidence suggest that the AMPK-mTOR pathway participated in the regulation of oxidative stress [22–24]. Wherefore, it is essential to investigate the effect of tributyrin on AMPK-mTOR signaling pathway. Our results revealed for the first time that tributyrin elevated the phosphorylation of AMPK and reduced phosphorylation of mTOR in intestine of pigs. These findings are consistent with previous results where SCFAs stimulate AMPK and inhibit mTOR in human colon cancer cells [57]. Similarly, Mollica et al. [58] found that in insulin-resistant obese mice, sodium butyrate increased liver AMPK activity, reduced ROS generation and improved mitochondrial function.
Conclusions
To summarize, the current experiment proved that dietary supplementation with tributyrin enhances intestinal mucosa architecture, promotes the intestinal growth, extenuates oxidative stress and modulates the AMPK-mTOR signal pathway of weaned pigs.
For more of the article, please click here.
Article made possible through the contribution of Caihong Hu et al.