Antimicrobial resistance on farms: a review including biosecurity and the potential role of disinfectants in resistance selection
Abstract: Resistance to therapeutic antimicrobial agents is recognized as a growing problem for both human and veterinary medicine, and the need to address the issue in both of these linked domains is a current priority in public policy. Efforts to limit antimicrobial resistance (AMR) on farms have so far focused on control of the supply and use of antimicrobial drugs, plus husbandry measures to reduce infectious disease. In the United Kingdom and some other countries, substantial progress has been made recently against targets on agricultural antimicrobial drug use. However, evidence suggests that resistant pathogenic and commensal bacteria can persist and spread within and between premises despite declining or zero antimicrobial drug use. Reasons for this are likely complex and varied but may include: bacterial adaptations to ameliorate fitness costs associated with maintenance and replication of resistance genes and associated proteins, horizontal transmission of genetic resistance determinants between bacteria, physical transfer of bacteria via movement (of animals, workers, and equipment), ineffective cleaning and disinfection, and co-selection of resistance to certain drugs by use of other antimicrobials, heavy metals, or biocides. Areas of particular concern for public health include extended-spectrum cephalosporinases and fluoroquinolone resistance among Enterobacteriaceae, livestock-associated methicillin-resistant Staphylococcus aureus, and the emergence of transmissible colistin resistance. Aspects of biosecurity have repeatedly been identified as risk factors for the presence of AMR on farm premises, but there are large gaps in our understanding of the most important risk factors and the most effective interventions. The present review aims to summarize the present state of knowledge in this area, from a European perspective.
Introduction
Scope and aims of the review
The principal focus of the review is the influence of biosecurity and related measures on antimicrobial resistance (AMR). Earlier review articles are used to establish the key background and conceptual material, while recent studies are used to illustrate contemporary examples. The evidence in this area arises from diverse investigations, for example, longitudinal and trace-back studies prompted by an observed AMR issue, formal sampling and risk-factor analyses of AMR on livestock units, and focused technical investigations into disinfectant effects or the effects of AMR on bacterial fitness and persistence. The review seeks to illuminate key ideas with detailed consideration of pertinent examples, rather than to provide comprehensive coverage of the field.
The problem of AMR
AMR among bacteria increases the morbidity and mortality associated with infectious disease. In the field of human medicine, this encompasses not just treatment of natural bacterial disease, but the control of infections attendant upon many other conditions and procedures. Recent estimates of human deaths attributable to bacterial AMR are around 33,000 annually in the European Union plus the European Economic Area (Cassini et al., 2019), and hundreds of thousands globally (Review on Antimicrobial Resistance, 2014, p. 18). Conserving the effectiveness of existing antimicrobial drugs is an urgent priority, as the capacity of pharmaceutical research and current finance models to develop and bring to market novel and affordable antibacterial drugs is in some doubt (Review on Antimicrobial Resistance, 2015, p. 42). The World Health Organization has highlighted eight pathogens of particular concern with respect to AMR, which include five bacterial species (Klebsiella pneumoniae, Escherichia coli, Staphylococcus aureus, Neisseria gonorrhoeae, and Mycobacterium tuberculosis) plus the agents of malaria, HIV/AIDS, and influenza (WHO, 2018).
In veterinary medicine the challenge of AMR to effective therapy of bacterial disease exists, but for livestock there are other considerations also. In competitive markets, narrow profit margins and high productivity mean that the effects ofinfectious disease can be highly significant financially as well as for health and welfare. In the 2000s the use of veterinary antimicrobials as growth promoters in the European Union was prohibited (European Parliament and European Council, 2003). However, antimicrobial drugs have continued to be used to counteract endemic or recurrent disease, often in prophylactic and group-medication modes (EFSA, 2011b; Hockenhull et al., 2017; Rhouma, Beaudry, Thériault, & Letellier, 2016). This can lead to poor or increasingly expensive disease control (such as with Brachyspira spp. in pigs (Karlsson, Aspán, Landén, & Franklin, 2004)), and it has also generated understandable alarm about the transfer of antimicrobial-resistant bacteria (or genetic elements conferring resistance) to humans and their microbiota, via food and other routes.
Current issues for AMR in livestock bacteria
Bacteria of concern. Current concerns about AMR transmission from livestock to humans are focused on resistant bacteria known to enter the food chain, or otherwise transmit to humans, which have zoonotic potential and/or which harbor mobile genetic elements (principally plasmids) encoding AMR. Zoonotic bacteria of concern are: Salmonella enterica, Campylobacter, and livestock-associated methicillin-resistant S. aureus (LA-MRSA). Organisms with less zoonotic potential, but which pose a threat to susceptible individuals and may carry mobile resistance determinants, include: E. coli, other members of the Enterobacteriaceae (particularly Klebsiella spp.) and Enterococcus spp.
Livestock-associated MRSA. LA-MRSA can colonize humans and may go on to cause antimicrobial-resistant disease in some circumstances (Becker, Ballhausen, Kahl, & Köck, 2017; Soavi et al., 2010). Incidence of LA-MRSA human colonization appears variable, according to prevailing local livestock activity and MRSA colonization (Feingold et al., 2012). The risk of severe disease appears to be lower than with MRSA of typical human subtypes. In a single-hospital study in Germany, admission screening at one hospital identified 25% of MRSA isolates to be of livestock-associated type, yet LA-MRSA amounted to only 7% of MRSA isolates from clinical sample submissions (Köck et al., 2011). The identification since 2005 of widespread LA-MRSA colonization of pig herds in many countries with intensive pig production (EFSA, 2009; Lassok & Tenhagen, 2013), plus in retail pork and other meats (Fox et al., 2017; Lassok & Tenhagen, 2013; Verhegghe et al., 2016), has led to a focus on characterization and control of this AMR organism, plus substantial interest in the media and among a general public familiar with MRSA from healthcare and com-
munity sources.
Third- and fourth-generation cephalosporin resistance. The extended-spectrum cephalosporin antibiotics are highly important for the treatment of serious human Salmonella, E. coli, and Klebsiella infections (WHO, 2016, p. 41). They readily select for resistance among Enterobacteriaceae in livestock, via genes of the extended-spectrum beta-lactamase (ESBL) or AmpC type. These resistances, whether plasmid-borne (ESBL, AmpC) or chromosomal (AmpC), have frequently been found in livestock and in derived foodstuffs (Liebana et al., 2013) and may often be found in association with other mobile resistances (Baron et al., 2018). There appears to be limited overlap between extended-spectrum cephalosporinase E. coli recovered from animals (or derived foods) and the general human population (Börjesson et al., 2016; Day et al., 2016; Nguyen et al., 2019; Wu et al., 2013). However, human–animal sharing of closely related strains or plasmid types has been observed, especially in farmers (Dorado-Garc´ ıa et al., 2018; van Hoek, Stalenhoef, van Duijkeren, & Franz, 2016), and consequently there is a particular focus around the licensing and use of these antibiotic classes in agriculture.
Carbapenem resistance. Resistance to this class of critically important human drugs has been detected among Enterobacteriaceae on pig farms in Germany, the United States, and China, plus in farmed and wild birds (Fischer et al., 2013; Köck et al., 2018; Kong et al., 2017; Mollenkopf et al., 2017; Roschanski et al., 2017). Carbapenems have never been licensed for veterinary use and resistance is a growing problem in human hospital and community medicine. Therefore, carbapenem resistance in livestock and wildlife is likely to have originated from human sources. Carbapenem resistance potentially can be amplified, spread, and re-presented to human populations by animals, especially as carbapenemases confer resistance to veterinary-licensed beta-lactam antimicrobials and many are encoded on transmissible plasmids.
Colistin resistance. Colistin, a polymyxin antimicrobial has been used to control gastroenteritis in groups of pigs typically at the postweaning stage (and to a lesser extent in poultry also) for decades, although in some non-European Union territories it may also still be used in a less targeted fashion as a growth promoter (Rhouma et al., 2016). In recent years colistin resistance has increased among pig Enterobacteriaceae isolates (Rhouma et al., 2016) and in the last two decades its importance in human medicine has grown, despite its potential for nephrotoxicity, as a treatment for increasingly intractable gram-negative multiresistant infections (Falagas & Rafailidis, 2008; WHO, 2016, p. 41). Recently, plasmid-borne resistance to colistin has been identified among some Enterobacteriaceae from human and farming sources (Rebelo et al., 2018; Rhouma et al., 2016), with organisms of the genus Moraxella being a suspected source of the mobilized mcr resistance genes (AbuOun et al., 2017). Furthermore, transmission between pigs and humans of E. coli carrying mobile mcr genes has been documented (Olaitan et al., 2015). Accordingly, polymyxins have been classed as critically important human antimicrobials (WHO, 2016, p. 41) and the use of colistin in animals is under scrutiny. Plasmid-borne mcr-1 colistin resistance has recently been reported from multidrug-resistant (MDR) E. coli and Salmonella isolates from pigs (United Kingdom) and poultry (Tunisia), underlining the additional threats of co-resistance and mobility posed by mcr-bearing organisms (Anjum et al., 2016; Grami et al., 2016; Maamar et al., 2018).
Multidrug resistance. MDR is defined as resistance to three or more classes of antimicrobial drugs; it is commonly seen among livestock isolates of E. coli (Cheney et al., 2015; Piccirillo et al., 2014) and S. enterica (European Centre for Disease Prevention and Control, European Food Safety Authority, & European Medicines Agency, 2017; Gomes-Neves et al., 2014; Mueller Doblies, Clouting, & Davies, 2013) in Europe. The occurrence of MDR in the latter (zoonotic) species is a focus of particular concern. MDR is also a common feature of LA-MRSA isolates (Lassok&Tenhagen,2013;Tenhagenetal.,2018;Verheggheetal., 2016).
Co-resistance. Concern about ESBL and fluoroquinolone (FQ) co-resistance (genetic linkage) in members of the Enterobacteraceae relates particularly to its potential transmissibility. Resistance to third- and fourth-generation cephalosporins is typically plasmid-borne, while FQ resistance characteristically follows from chromosomal mutations (European Centre for Disease Prevention and Control, European Food Safety Authority, & European Medicines Agency, 2017). However, there are plasmid-encoded genes and gene variants that can promote the development of clinical FQ resistance, or augment it. These have recently been found co-located on ESBL plasmids in livestock or meat-derived E. coli (Ghodousi, Bonura, Di Noto, & Mammina, 2015; Zurfluh, Cernela, & Stephan, 2015) and Salmonella (Jiang et al., 2014) strains.
Drivers for AMR
Antimicrobial usage. There is ample epidemiological evidence that antibiotic usage in humans is causally related to the emergence and increased degree of AMR in treated populations (Hawkey & Jones, 2009; Scientific Advisory Group on Antimicrobials of the Committee for Medicinal Products for Veterinary Use, 2009). Observational studies and risk factor analyses of commensal and pathogenic bacteria among veterinary species at farm and national levels (Bosman, Wagenaar, Stegeman, Vernooij, & Mevius, 2014; Dierikx, van der Goot, Smith, Kant, & Mevius, 2013; Dohmen et al., 2017; Prásek, Sperling, Lobová, Smola, & Cızek, 2014; Scientific Advisory Group on Antimicrobials of the Committee for Medicinal Products for Veterinary Use, 2009; Taylor, CliftonHadley, Wales, Ridley, & Davies, 2009; Taylor, Wales, Ridley, & Davies, 2016) also provide convincing evidence of such an association between antimicrobial usage and resistance. Well-established mechanisms for this include: mutations that affect antibiotic target sites or increase the expression or spectrum of existing resistance genes, and selection for horizontal spread of resistance genes (Scientific Advisory Group on Antimicrobials of the Committee for Medicinal Products for Veterinary Use, 2009).
Investigations of ESBL and AmpC-mediated β-lactam and cephalosporin resistance among poultry pointed to the use of cephalosporins in hatcheries (administered in ovo or at hatching) as a likely driver for colonization of birds by E. coli (and, in one study, Salmonella) that were cephalosporin resistant (Dutil et al., 2010; Persoons et al., 2011), with one study showing hatchery use to be highly significant in a multivariable regression model for the flock-level prevalence of cephalosporin resistant E. coli (Baron et al., 2014). More broadly, recent rapid expansion of ESBL-type resistance among nonclonal isolates of E. coli from livestock strongly suggests horizontal transmission as the dominant mode of resistance acquisition (Scientific Advisory Group on Antimicrobials of the Committee for Medicinal Products for Veterinary Use, 2009).
A further contemporary example of antibiotic usage driving resistance, this time mediated by chromosomal point mutations, is the progressive increase over the last two to three decades in the frequency and multiple nature of AMR of Brachyspira spp. isolates from pigs, particularly Brachyspira hyodysenteriae. This has occurred in the face of antimicrobial drug use (lincosamide, macrolide, and pleuromutilin classes), primarily against swine dysentery (Mahu et al., 2017; Prásek et al., 2014; Rugna et al., 2015; Sperling, Smola, & Cızek, 2011).
Spread of resistance. One effect of antimicrobial drug use on farms may be the increased spread of mobile AMR genes by mechanisms (probably including global stress responses) that enhance the frequency of conjugative transfer between bacteria (Gillings, 2016). Such effects differ between antimicrobial drugs, and also occur with sublethal exposure to certain disinfectants (Jutkina, Marathe, Flach, & Larsson, 2018). However, the balance of the influences of antimicrobial compounds (in the broadest sense) on the horizontal transfer of AMR on farms is poorly understood, as it may include inhibition as well as promotion of gene transfer. Work on antimicrobials used as growth promoters, plus some (including chlortetracycline and tylosin) used for prophylaxis and therapy, showed considerable inhibition of plasmid transfer into E. coli at normal in-feed concentrations (Mathers et al., 2004), while chlorhexidine and some quaternary ammonium compounds (QACs) (applied at or just below their minimum inhibitory concentrations [MICs]) also reduced plasmid transfer (Al-Masaudi, Day, & Russell, 1991). Some antibacterial compounds, such as certain unsaturated fatty acids, have a specific antiplasmid-transfer effect (Getino et al., 2015).
In addition to genetic transfers, once antimicrobial-resistant bacteria are present on livestock units or in associated enterprises (such as hatcheries, slaughterhouses, livestock markets/collection centers, feed mills, and transportation), there exists the possibility for physical transfer of resistant organisms elsewhere, either within or between premises. This may occur without any selective antimicrobial pressure. The following summaries of field studies illustrate the spatial transfer of AMR.
The presence of AmpC-type cephalosporin-resistant E. coli on dairy farms in Ohio was associated with third-generation cephalosporin (ceftiofur) use in the herd (univariable logistic regression, odds ratio [OR] 25, P < 0.01), whereas isolation from individual adult animals was not associated with recent administration of such a drug (Tragesser, Wittum, Funk, Winokur, & Rajala-Schultz, 2006). This suggests that resistant strains may readily disseminate from treated animals to others within an epidemiological group. Similarly, in a multivariable regression model the presence of FQ resistance among E. coli from postpartum cows was a risk factor for the presence of FQ-resistant E. coli in calves on the same premises (P < 0.001; Duse et al., 2015). Apparent spread between species was observed using molecular and resistance typing of LA-MRSA, showing colonization of broilers, rats, dogs, cats, and farmers by strains highly similar to those in pigs on the same premises (Pletinckx et al., 2013). In a pattern already established for other contagious mastitis pathogens, poor hygiene at milking was observed to be associated with LA-MRSA spread between dairy cows (Locatelli et al., 2017).
Several studies examining the detection and/or subtyping of AMR bacteria have yielded evidence of the physical transfer of AMR between premises and between generations. This has been documented for LA-MRSA in pigs and cattle (Grøntvedt et al., 2016; Locatelli et al., 2017; Pletinckx et al., 2013), for cephalosporin-resistant E. coli in broilers (with hatcheries appearing to act as source and/or conduit in many cases) (Baron et al., 2018; Dierikx et al., 2013), and for FQ-resistant Campylobacter coli in broilers (Idris et al., 2006). An alternative (metagenomic) approach has yielded evidence of associations between the frequencies of certain AMR genes in feces, comparing breeding sows and their derived finishing herds on separate premises (Birkegård et al., 2018).
Other studies have noted the occurrence of clonal and multiresistant subtypes of B. hyodysenteriae (some of them novel) on numerous apparently unrelated pig farms in Italy (Rugna et al., 2015) and Belgium (Mahu et al., 2017). This has been interpreted as evidence of spread by physical transfer, although whether by animals, workers, or equipment is unclear. Clonally related ESBL-bearing E. coli have been noted on apparently unconnected Dutch veal calf premises (Hordijk et al., 2013).
Investigations involving the international movement of humans and of breeding animals have, similarly, provided evidence of AMR transfer across national boundaries (Dierikx et al., 2013; Hawkey & Jones, 2009). A European Union summary report in 2011 concluded that there was evidence of a spread of cephalosporin-resistant E. coli from imported breeding stock into Swedish broiler production, and that the occurrence of such organisms in the various levels of the Dutch poultry production chain was the result of vertical transmission, local recirculation, and selection (EFSA, 2011b).
Persistence. Once present on a farm, antimicrobial-resistant bacteria may be isolated repeatedly from subsequent groups of animals, even in the face of short production cycles, frequent cleaning and disinfection (C&D), and little external selection pressure. In a Norwegian study of broilers, cephalosporin-resistant E. coli (believed to have been imported with breeding stock) all shared a common resistance phenotype and genotype (AmpC, CMY-2) and persisted in production flocks at a higher frequency than among parent flocks, in the absence of antimicrobial use (Mo, Kristoffersen, Sunde, Nodtvedt, & Norstrom, 2016). In the same study, multivariable modeling (further detail in Risk Factors for AMR in Commensal Bacteria section) indicated the risk of a flock being positive to be significantly and substantially elevated when the previous flock was affected, but reduced when effective floor disinfection was always performed between flocks, indicating carryover between flocks to be a likely factor in repeated isolations.
In surveys conducted on numerous pig and poultry units, the proportion of premises yielding FQ-resistant E. coli and Campylobacter was similar regardless of whether FQ had been used recently or more than a year previously (Taylor, Clifton-Hadley, Wales, Ridley, & Davies, 2009, 2016). Furthermore, in Australia (a country where FQ had never been licensed for livestock), closely related E. coli strains carrying chromosomal FQ-resistance mutations plus multiple plasmid-mediated AMRs were isolated commonly from poultry carcasses (Ingram, Rogers ,Sidjabat ,Gibson, & Inglis, 2013). It was hypothesized in the latter case that persistence of FQ resistance could be driven by co-selection of MDR among E. coli strains by non-FQ antimicrobials, although the origin of the chromosomal FQ-resistance mutations remains obscure. Co-selection by a heavy metal (zinc) is a possible mechanism for the maintenance of multidrug resistant E. coli in piglets around weaning, an association suggested by a recent experimental study (Ciesinski et al., 2018). Finally, a dairy farm study showed that a LA-MRSA strain persisted on a unit for at least two years, albeit presumably under conditions of routine antimicrobial use (Locatelli et al., 2017).
Initiatives on control of resistance
Current reports and strategies on tackling AMR in agriculture (Anon., 2013, pp. 34–37; Review on Antimicrobial Resistance, 2016, p. 81; RUMA, 2014; Speksnijder, Mevius, Bruschke, & Wagenaar, 2015) emphasize measures to reduce antimicrobial drug use, including: restrictions on drug licensing and prescribing, surveillance of drug use, and husbandry and biosecurity measures to prevent or control infectious disease.
Opinions vary on the financial and welfare costs and benefits of reducing agricultural antimicrobial drug use. Some studies on pig units observe a positive correlation between lower antimicrobial use and higher measures of biosecurity anda nimal health (Isomura, Matsuda, & Sugiura, 2018; Postma et al., 2016). However, this may represent something of a "virtuous circle" and, by contrast, no link between antimicrobial use and economic or production performance was seen in a study of turkey production (Chauvin et al., 2005). The effects of a forced or nontargeted reduction in antimicrobial use may be less benign, especially in other sectors (Lhermie, Tauer, & Gröhn, 2018). Expert opinion in Europe shows wide variation in the perceived scope for antimicrobial drug use reduction, influenced by differences between countries, sectors and personal experience (Carmo et al., 2018). There was, however, consensus in the last study on the potential value of improved biosecurity, veterinarian education, and the use of treatment plans to facilitate such reductions.
Initiatives by the European Union aimed at protecting human populations from AMR originating from agriculture are typified by joint interagency antimicrobial consumption and resistance analysis (JIACRA) reports and the antimicrobial advice ad hoc expert group (AMEG). The most recent JIACRA report assessed data from European Union livestock, performing multiple com-parisons of antimicrobial drug use with susceptibility of indicator bacteria (European Centre for Disease Prevention and Control, European Food Safety Authority, & European Medicines Agency, 2017). There was mixed evidence of correlations, and signifi-cant relationships were not identified in many pairings of drug and bacterial species, including cases involving extended-spectrum cephalosporins and FQ. Nonetheless, all such initiatives currently lack substantial consideration of mitigating the risks of AMR be-ing introduced to livestock-related premises via animals, workers, equipment, and feed, or of AMR persisting in the absence of external selective pressures.
Studies on AMR on Farms
A number of investigations have examined the occurrence of AMR on farms in relation to other factors. These have generally been qualitative determinations, using antibiotic-containing selective media to isolate organisms with reduced susceptibility to the agents in question. Typically, these are then subjected to further characterization. A few investigations (Boulianne et al., 2016; Duse et al., 2015; Schuppers et al., 2005) have not used selective resistance-screening media for initial isolation, or have attempted a quantitative determination by using selective and nonselective media. Figure 1 provides a schematic overview of principal findings in this area.
Risk factors for AMR in pathogenic bacteria
There exist a few formal risk factor analyses investigating the presence of AMR in zoonotic pathogens on farms. In these, summarized below, themes of antimicrobial use, biosecurity, and hygiene are dominant.
Tayloretal. (2009) examined FQ-resistant thermophilic Campylobacter on U.K. finisher pig farms. Among 108 pig units, any previous use of FQ was, as expected, a positive risk factor (OR 8.3), yet 54% of farms with no reported use of FQ also yielded FQ-resistant isolates. Multivariable regression provided four additional highly plausible risk factors at or near conventional significance, which related to external biosecurity (buying-in of grower pigs [OR 4.3], visitor biosecurity regulations [OR 0.33]) or on-farm hygiene.
A related investigation of 89 broiler and turkey farms (Taylor et al., 2016) similarly found FQ use was a clear risk factor (OR 14.7), but FQ resistance was present on many units without reported use of such antimicrobials. Of the additional risk factors at or near conventional significance, wild bird access to poultry houses was a positive risk (OR 10.3, P < 0.017) while three hygiene measures (provision of a face mask to staff, all detailed areas being dusted, and feed hoppers being cleaned and disinfected) were apparently protective. These negative factors were variously and positively correlated with other hygiene practices: wet cleaning of all detailed areas with disposal of wash water off-site, C&D of ante rooms, provision of hat and gloves, hand sanitizer, and a toilet. Larger and independently run farms were also at lower risk.
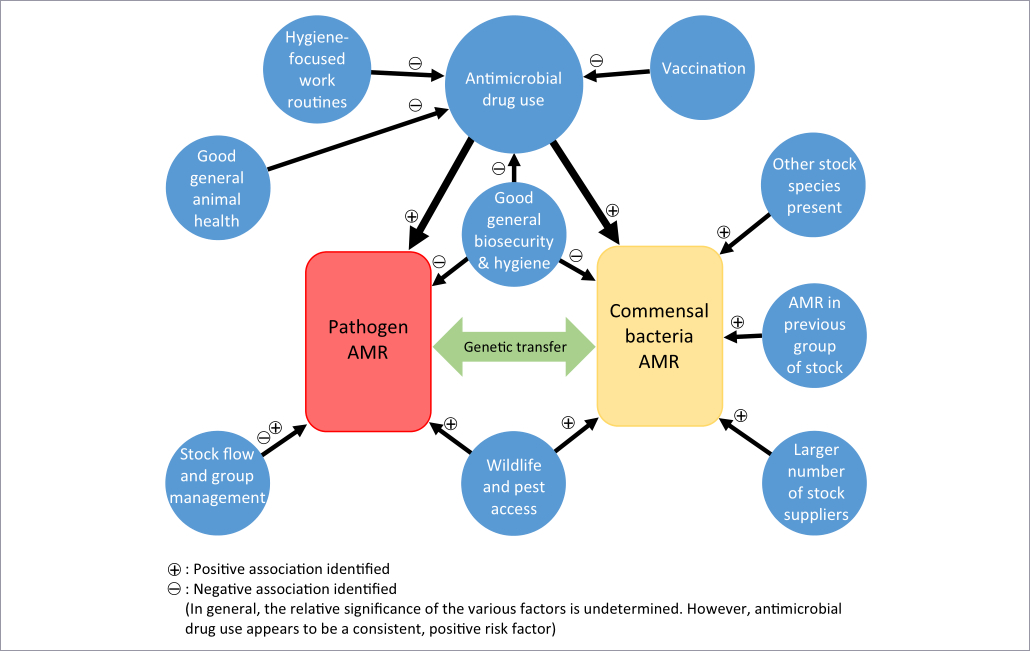
Figure 1 –Schematic diagram of some risk relationships between antimicrobial resistance (AMR) and farm factors, as identified in field studies of livestock-associated bacteria.
Multivariable regression using C. coli resistance data from 64 Swiss pig farms identified partially all-in-all-out production as a risk for MDR compared with strict all-in-all-out systems (OR 36.6, P < 0.001; Schuppers et al., 2005). Positive risk factors associated with multiple or individual antibiotic resistances commonly were related to pig health or injury. Antimicrobial drug administration was not included in models, owing to unreliable farm records.
Modeling performed using data from a European Union-wide environmental survey of pig units conducted in 2008 indicated that the risk of a holding being LA-MRSA-positive increased with the number of breeding pigs on the premises, for both production and breeding holdings (EFSA, 2010). Furthermore, there were strong correlations between the country-level prevalence of MRSA-positive breeding (genetic or multiplier) pig holdings and both the volume of imported breeding pigs (correlation coefficient = 0.6, P < 0.002) and the prevalence of MRSA-positive production holdings (correlation coefficient = 0.76, P < 0.0001). These findings strongly suggest that MRSA risk is associated with the movement of pigs, both internationally and locally.
Risk factors for AMR in commensal bacteria
E. coli has been the main bacterial species of choice for studies monitoring and analyzing AMR in livestock populations. It is ubiquitous, known to be involved in some diseases (particularly of young stock) and in the hosting and exchange of resistance determinants, and is relatively easily isolated and analyzed, including for gene transfer studies. In common with the previously described analyses of AMR pathogens, biosecurity- and hygiene-related factors emerge repeatedly as significant risk factors for AMR in the field studies summarized below.
In Norwegian commercial broiler production, and in the absence of antimicrobial use, a multivariable model revealed associations between the presence in a flock of cephalosporin-resistant E. coli (AmpC-type) and: the previous flock being similarly positive (OR 12.7, P < 0.001), more parent flocks contributing to the production flock (3 versus 1 parent flock: OR 6.3, P = 0.01), chick delivery personnel being allowed access to the broiler house (OR 9.3, P = 0.01), and consistent floor disinfection between flocks (negative association; OR 0.1, P = 0.01) (Mo et al., 2016). Among Canadian broilers, the administration of ceftiofur (thirdgeneration cephalosporin) to a flock at the hatchery was associated with a higher proportion of ceftiofur-resistant E. coli isolated from the flock at slaughter, in a bivariate analysis (Boulianne et al., 2016). Furthermore, the same study observed that rearing of turkeys on litter previously used by a broiler chicken flock was associated with carriage of ceftiofur-resistant E. coli in the turkey flock (univariable logistic regression). A multivariable analysis of E. coli in U.K. turkey production (where cephalosporins are not licensed) found the presence of cephalosporin resistance in fattening flocks to be associated (P ? 0.02) with certain external biosecurity factors (nearby pig units [OR 2.6], staff working with other livestock [OR 2.8]), while some flock separation measures (dedicated gloves and house partitions) appeared protective (P ? 0.09; Jones et al., 2013).
The same U.K. study (Jones et al., 2013) also considered FQ (ciprofloxacin) resistance. For turkey fattening flocks, any antimicrobial use in the flock or evidence of mouse activity was weakly associated with an increased risk of resistant E. coli being present (P ? 0.1), while routine disinfection at depopulation and partitioning of houses appeared protective (OR 0.38, P = 0.035). Among breeding flocks (albeit with a small data set), risk was increased in association with flocks that were larger or that had used FQ in the previous year, whereas frequently changed foot dips were apparently protective. Another U.K. study, of the presence of ciprofloxacin-resistant E. coli in turkeys and broiler production flocks (Taylor et al., 2016), identified certain biosecurity features as risk factors, that is, public perimeter path (positive; OR 4.1, P = 0.019) and perimeter fence (protective; OR 0.27, P = 0.014). The use of FQ and single-handed operation were also positive risk factors.
In a study of Dutch pig production using multivariable regression (Dohmen et al., 2017), isolation of ESBL-positive E. coli from pooled rectal swabs from 36 farrowing herds (in the context of markedly reduced cephalosporin use in previous years) was associated with use of cephalosporins up to 6 months previously (OR 72, P = 0.001) but considerably less likely if there was a oneway entry room to change into dedicated clothing and footwear (hygiene lock) at the farm entrance (OR 0.06, P < 0.001). The presence of other livestock (specifically goats) on the farm was also a positive risk factor, which may relate to lower biosecurity standards. For the presence of FQ (ciprofloxacin)-resistant E. coli on U.K. finisher pig units, use of FQ was the dominant risk factor (Taylor et al., 2009). However, other factors at or near significance in the multivariable model were: a nearby pig farm (OR 3.5, P = 0.076) or warmer season (summer versus autumn/winter: OR 12.7, P < 0.005), and visitors required to have been free of pig contact for two days (protective; OR 0.4, P < 0.1). Having a poultry farm nearby was also protective in the same study, possibly owing to a "dilution" effect on exogenous pig-derived E. coli strains.
FQ resistance in rectally obtained E. coli was examined in a multivariable regression study of postpartum cows and preweaned calves on Swedish dairy farms with historical use of FQ, as discussed in Current Issues for AMR in Livestock Bacteria section (Duse et al., 2015). Using earlier data, units with high (>10%) and zero percentages of FQ-resistant E. coli were included. In addition to the already-described association between FQ-resistant E. coli in any sampled postpartum cow and calves on the same unit, there were associations between an increased percentage of FQ-resistant E. coli in a calf sample and: calf age (there was a striking reduction from around 18 days of age), any use of FQ in cattle within the previous four months (P < 0.001), and feeding milk from cows treated either with antimicrobials generally or with FQ specifically (P ? 0.006). For adult postpartum cows, the risk of isolating resistant E. coli was raised when they were group-housed or in open herds (OR 7.9 and 6.9, respectively, P = 0.02), or when the farm shared transport with other farms (OR 3.6, P = 0.002) or had a below-average hygiene score (OR 3.6, P = 0.021).
Cross-sectional studies on dairy farms have identified the use of third- and fourth-generation cephalosporins as risk factors for the presence of E. coli expressing CTX-M-type ESBL (Snow et al., 2012) or ESBL/AmpC resistance (Gonggrijp et al., 2016; Tragesser et al., 2006). In multivariable regression models, CTX-M presence was negatively associated with high biosecurity (closed herds compared with un-quarantined open herds; OR 0.05, P = 0.004) and superior calf feeding equipment hygiene (65 U.K. herds, Snow et al., 2012). The presence of ESBL/AmpC correlated with treatment of all mastitis cases with antimicrobial drugs (OR 3.9, P = 0.02) and increasing proportion of calves treated with antimicrobials (86 Dutch herds, Gonggrijp et al., 2016). In the latter study, various features of biosecurity were not identified as risk factors, but much of the variation remained unexplained in the final model and follow-up testing 6 months later showed that many farms had changed status, possibly owing to issues with sensitivity of detection.
Among gram-positive bacteria, Enterococcus species are commonly examined as sentinels for AMR, given that they are considered part of the normal livestock intestinal microbiota, they express diverse patterns of AMR, and they frequently pose an AMR problem in human medicine. However, reports examining risk factors for AMR among enterococci from livestock are sparse. Significant associations, using bivariate analyses, were found in Canadian poultry between the individual-level prevalence of resistance to specific antimicrobial drugs among enterococci at slaughter and use of those drugs at the hatchery (gentamicin, turkeys, P = 0.04) or during rearing (tylosin, broiler chickens; bacitracin, turkeys; bacitracin, broilers; P < 0.01 in all cases) (Boulianne et al., 2016).
Risk factors for the use of antimicrobial drugs on farms
Biosecurity and hygiene also feature prominently in studies examining differences between farms' use of antimicrobial drugs. This lends weight to the reasonable perception that the AMR, antimicrobial drug use, biosecurity and disease control through hygiene are all interrelated. Relevant publications are outlined below.
Correlations between antimicrobial use and other herd-level parameters were examined, in a multivariable regression model, on 227 farrow-to-finish farms in Belgium, France, Germany, and Sweden (Postma et al., 2016). Lower antimicrobial use, on a standardized measure, was associated (P < 0.01) with better external biosecurity scores (using the Biocheck.UGent tool) and vaccination against more pathogens, but not with either daily live-weight gain or mortality.
A related study divided the same 227 farms by production and antimicrobial use, using country-specific median values for reference (Collineau et al., 2017). Farms with above-average weaners per sow per year and below-average standardized antimicrobial usage (plus no FQ or third- and fourth-generation cephalosporins) were not significantly different from all the other farms with respect to herd characteristics, but they had significantly better biosecurity and hygiene markers (local density of pig units [OR 3.8, P = 0.012], and order of work on-farm from more disease-susceptible young pigs to older pigs [OR 2.4, P = 0.044]) and disease markers (less gastrointestinal disease and less respiratory disease among suckling pigs and fatteners, respectively). In areas with higher pig unit density, work routines and equipment deployment that progressed systematically from younger to older stock were both significantly correlated with the high production/low antimicrobial use farm subset.
In turkey production in France, the overall intensity of antimicrobial drug use was found to be similar between successive flocks on any given premises, but not to be significantly associated with the economic or technical performance of a unit (Chauvin et al., 2005). Multivariable regression analyses identified lower antimicrobial consumption (that is, in the bottom third of the study farms on a standardized measure of antimicrobial use) to be associated with the external biosecurity measure "changing of shoes and clothes on entry to premises" (OR 2.7, P = 0.02). The same risk factor was negatively associated with farms in the top third of the study population for antimicrobial use.
Farm studies on the prevalence of AMR genes
Not much work has been published in this area. A study by Birkegård et al. (2018) examined the copy number of certain AMR genes in feces from sows, and from their finisher offspring reared on different premises. Both of the macrolide (ermB, ermF) and one of the tetracycline (tetO) genes were significantly correlated (P values between 0.02 and 0.04, correlation coefficients between 0.33 and 0.47) at a farm level between dams and their offspring, suggesting transfer of AMR between the groups of animals. In another study, high-level supplementation with zinc oxide (used to manage postweaning piglet diarrhea), was associated (bivariate analysis, P ? 0.05) with increased tetracycline (tetA) and sulfonamide (sul1) resistance gene copy numbers in feces of weaners after two to three weeks (Vahjen, Pietruszy´ nska, Starke, & Zentek, 2015).
Intervention study on antimicrobial use
Farmers from 61 Belgian pig units were recruited and audited, then biosecurity, preventive medicine, and antimicrobial use recommendations were made for each farm (Postma, Vanderhaeghen, Sarrazin, Maes, & Dewulf, 2017). Implementation plus performance and antimicrobial use was assessed eight months later. Overall, significant improvements in biosecurity scores were seen in 11 of 14 measures (paired data, P ? 0.05), albeit in areas perceived to be easier and cheaper to implement, and there was a significant reduction in antimicrobial use for disease prevention in young stock up to weaning (paired data, 0.01 < P ? 0.02). Nonsignificant reductions were seen in preventative antimicrobial use in older stock, and in all therapeutic use. Allied to this were small but significant improvements in production parameters: weaners per sow per year, daily live-weight gain, mortality, and feed conversion.
Transmission and Persistence of Antimicrobial-Resistant Organisms
Table 1 summarizes factors, discussed in this and subsequent sections, on the persistence of AMR on farms. Some factors are interrelated such as: animal management systems, opportunities for hygiene controls, and C&D. The transmission and movement of AMR, rather than the generation or selection of AMR in situ by antimicrobial agents, has been observed in several field studies, some of which are outlined in the following section. Identified or postulated routes of transmission included: importation of breeding stock with subsequent transfer down a production pyramid, repeated introductions into groups of young animals from a persistent source (hatchery, adult cattle herd), and movement between premises or areas linked by personnel, stock, vehicles, or wind. Where resistant bacteria appeared not to be closely related, likely sources of AMR included intensive use of antimicrobial drugs, or transmissible genetic elements (plasmids); both of these were associated with hatcheries.
A study by Dierikx et al. (2013) examined cephalosporin resistance in E. coli within the highly structured multilevel operation of the Dutch broiler sector. Some details of the findings are presented, with a summary in the following paragraph. AmpC-type resistance was found in newly hatched grand-parent stock imported from the United Kingdom and Ireland at mean individualprevalence values of 23% and 44% for birds hatched in the United Kingdom ("breed A") and the Netherlands ("breed B"), respectively. The U.K.-hatched birds yielded ESBL and AmpC resistance in varying proportions of birds after FQ and penicillin treatments: at 31 weeks of age, sample prevalence per house was between 2% and 27%. Meconium taken from parent-level chicks yielded resistant E. coli: 0.31% of "breed A" samples were ESBL-positive and 5.8% of "breed B" samples were AmpC-positive. AmpC- and ESBL-expressing E. coli were also isolated from "breed A" broiler chick meconium samples (1.9%). In production houses the proportion of broilers yielding AmpC/ESBL-expressing E. coli rose from 20% or less on delivery to a sustained 100% at three to five weeks of age. Environmental samples taken from parent stock and broiler hatcheries, and also from floors of broiler houses before placement, were also positive for AmpC and ESBL phenotype E. coli in varying proportions. Cephalosporin-resistant isolates from all levels of the breeding pyramid also showed co-resistance to non-beta-lactam antimicrobials.
Thus, cephalosporin resistance was present from hatching of imported breeding stock. It was detected in increased variety, and in combination with other AMR, at hatching in lower levels of the breeding pyramid and in some hatchery environment samples, and within production flocks it was found within cleaned houses and at high prevalence among birds. Possible reasons for the higher prevalence within broiler flocks compared with grandparent flocks include relatively more frequent antibiotic use and lower standards of internal biosecurity in broiler production flocks.
Another study examined E. coli and their extended-spectrum cephalosporin resistance plasmids from serial sampling of French free-range broiler flocks derived from two hatcheries (Baron et al., 2018). Cephalosporin-resistant strains showed varied pulsed-field gel electrophoresis subtypes, even from the same hatchery. ESBL resistance was borne on plasmids bearing other resistances also, and a subset of these from flocks from both hatcheries were highly similar to each other, and to published sequences of plasmids from Dutch, Swiss, and American isolates. AmpC (CMY-2) resistance was only found in flocks from one of the two hatcheries, on highly similar plasmids in varied E. coli subtypes. MDR was common but did not appear to be encoded on the AmpC plasmids. The occurrence of extended-spectrum cephalosporin resistance among diverse E. coli, often with other AMRs and sharing highly similar resistance plasmids, suggests that the ESBL/AmpC phenotypes may often be acquired by horizontal transmission. Furthermore, they may be maintained and selectedfor by use of non-beta-lactam antimicrobials.
A longitudinal study of quinolone-resistant E. coli from Norwegian livestock surveillance showed a stable and low frequency of resistant isolates over the last decade, except from broiler birds where the percentage of resistant isolates spiked in 2009 and climbed steadily between 2011 and 2016 (Kaspersen et al., 2018). This correlates with similarly high quinolone resistance among E. coli from Swedish broilers, despite little or no quinolone or FQ exposure in the respective industries. This pattern may be a consequence, in both countries, of resistance carried by grandparen tbirds imported from the United Kingdom and transmitted (at least partly) vertically (Wang, Pors, Olsen, & Bojesen, 2018) or pseudo-vertically through hatcheries (Börjesson, Guillard, Landén, Bengtsson, & Nilsson, 2016).
A collection of pathogenic E. coli isolated from colibacillosis cases in Italy between 2008 and 2010 (when third-generation cephalosporins were still used prophylactically in day-old chicks) showed diverse ESBL or AmpC-bearing plasmids on a background of multiresistance, in 8% of isolates (Niero et al., 2018), consistent with multiple introductions or selection events. A similar but far more marked picture of extended-spectrum cephalosporin resistance plus multiple other resistances was seen in E. coli isolates from Egyptian hatcheries (Osman et al., 2018).

A study of enteric gram-negative bacteria (mostly E. coli) shed by suckler calves in the United States revealed that over 90% of calves were colonized by ESBL-expressing organisms in their first year, despite no use of subtherapeutic antimicrobials in the herd and no prior use of therapeutic antimicrobials in the calves (Mir et al., 2018). All ESBL isolates examined were multidrug resistant. The prevalence of shedding peaked at around 74% of individuals, among 6- to 9-month-old calves, but was much lower (around 6%) in older calves, up to 12 months of age. Age and season were correlated, so it was not possible to observe which factor had more influence on the decline in prevalence.
An interesting study on potential airborne spread of AMR by E. coli was reported in brief by Chinese workers. E. coli with AMR profiles matching those on rabbit and poultry farms were isolated from air samplers placed up to 200 m downwind of the units (Yao,Gao, Chai, Cai, & Duan, 2007). No E. coli were isolated from 100 m upwind of the units.
LA-MRSA has its core resistance determinants encoded chromosomally, and therefore horizontal spread of resistance between bacteria has not been a feature of this potential zoonosis. The extent and spread of LA-MRSA colonization within farms was investigated in a cross-sectional sampling study of six Belgian pig farms known to harbor the organism, two of which also had broiler poultry and two others had dairy units (Pletinckx et al., 2013). Subtyping (spa, SCCmec, multi-locus sequencing and AMR phenotype) showed that within a farm identical or highly similar MRSA subtypes were found among pigs, their environment, broilers, and rodents. Similar subtypes were also isolated from dogs, cats, goats, and farmers. By contrast, cattle, visitors (vets), and incoming gilts harbored substantially different subtypes.
Local spread between farms was examined in a cross-sectional study conducted on two separate MRSA-positive Italian dairy farms, each with a nearby pig unit sharing some access and, in one case, a farm worker (Locatelli et al., 2017). Environmental and animal sampling revealed that some MRSA subtypes were present on both the pig and dairy units within each pair of premises, suggesting local spread or a common local source. Some subtypes found on one dairy farm were highly similar to those isolated from the same premises two years previously.
In Norway, and in the context of a low prevalence of LA-MRSA-affected farms, farm workers or visitors from other European countries were considered to be a likely route of introduction to pig premises, based upon epidemiological and subtyping data (Grøntvedt et al., 2016). Further epidemiological evidence implicated the movement of animals, shared personnel, or pig transport vehicles as likely mechanisms for the spread of MRSA between Norwegian farms.
For Brachyspira spp., another pathogen like LA-MRSA with chromosomally determined antibiotic resistance, patterns of resistance between linked farms have been discerned in the United States (Mirajkar, Davies, & Gebhart, 2016) and in Europe (ˇSperling et al., 2011). Furthermore, clonal spread of tiamulin-resistant B. hyodysenteriae between farms has been identified in the United Kingdom (Karlsson et al., 2004). Evidence of Brachyspira spp. in farmed and free-living wild boars illustrates the potential for wild pig vectors to pose a biosecurity challenge for the transfer of disease and of AMR.
Biosecurity
Definition and principles
Biosecurity in the context of livestock production may be defined as all measures taken to prevent both the introduction and the spread of infectious (and parasitic) agents on the farm (Dewulf, 2014). It can broadly be divided into "external" biosecurity, focused on preventing introduction of infectious agents, and "internal" biosecurity, focused on preventing their spread within an individual farm, and hopefully their further dissemination via farm wastes. "Infectious agents" include both disease-causing entities, and those that are part of a "normal" biological system, that nonetheless potentially carry undesirable traits such as antimicrobial or anthelmintic resistance.
External biosecurity addresses the risk of entry to premises of infectious agent carriers, for example, other livestock, wildlife, companion animals, humans, machinery, and vehicles. It also aims to prevent contact between farm livestock and outside risks (such as neighboring livestock), and includes measures to ensure that necessary access (for example, feed, workers, replacement stock, and certain vehicles) minimizes the risk of introducing infectious agents. Internal biosecurity is closely linked to hygiene and preventive medicine. It encompasses a host of husbandry measures, disease prevention and control, logical working practices and sequences, C&D procedures and competencies, waste management, and so on. These are all aimed at keeping infectious disease to a minimum, treating disease adequately while protecting healthy and vulnerable stock from infectious agents and infestations, separating epidemiological groups, and providing opportunities for clean breaks within the cycles of production and occupation.
Relevance for control of AMR in bacteria
The practice of biosecurity is necessarily multimodal and varies according to livestock sector and local conditions. Consequently, studying and interpreting associations between biosecurity and other factors (which might in practice be linked with biosecurity policies) can be challenging, especially if conducted on diverse premises. Measuring biosecurity as an independent variable can be approached by integrating a standardized set of quantitative or semiquantitative metrics into one or more overall scores (Ghent University, n.d.), or by developing models that use certain biosecurity elements to represent clusters of correlated measures (Taylor et al., 2009). Assessment of biosecurity can be done on the ground by visiting investigators (which could bias study populations away from those unwilling to allow access), or at a distance using producers' declarations of their own practices. Whatever the approach, investigators need to be alert to local features that may represent unforeseen biosecurity hazards.
Link to intensity of antimicrobial drug use. The application of consistently high standards of all-round biosecurity has beneficial effects for AMR control, not only by preventing AMR being brought onto premises but also by assisting to reduce the need for antimicrobials to treat bacterial disease, whether primary (for example, swine dysentery) or secondary infections associated with viral disease or insanitary conditions. Antimicrobial drug use is a primary driver for the development and maintenance of AMR on premises, and characteristics of pig farms that have been associated with lower antimicrobial use in observational (not interventional) studies include superior biosecurity and hygiene practices (Risk Factors for the Use of Antimicrobial Drugs on Farms section). By contrast, a Swedish risk factor study on pig farms did not find a significant association between biosecurity and antimicrobial use (Backhans, Sjölund, Lindberg, & Emanuelson, 2016), although the authors suggested that high prevailing biosecurity or individual disease outbreaks may have obscured such an association.
Links to observed AMR. Examples of specific biosecurity measures that have been associated with a lower degree of AMR on farms have been discussed under risk factor studies in Risk Factors for AMR in Pathogenic Bacteria and Risk Factors for AMR in Commensal Bacteria sections. They include: a hygiene lock at the farm entrance for ESBL in pigs (Dohmen et al., 2017), a restriction on visitors with recent pig exposure for FQ-resistant E. coli and Campylobacter in pigs (Taylor et al., 2009), restrictions on external chick delivery personnel entering growing houses for AmpC resistance in broiler E. coli (Mo et al., 2016), a closed herd for FQ-resistant (Duse et al., 2015) or ESBL (Snow et al., 2012) E. coli in dairy cattle, and staff not working with other livestock for cephalosporin resistance in turkey E. coli (Jones et al., 2013). Despite the apparent importance of biosecurity in livestock antimicrobial use and AMR, it is notable how very few published formal risk factor studies there are that examine such relationships.
Control of possible human-to-livestock transfers. While the current public focus is on the transfer of AMR from livestock to humans, there is much yet to be learned about possible transfer in the other direction (Rogers, Shaffer, Langen, Jahne, & Welsh, 2018) that has a bearing on biosecurity practices. Such transfers may occur via birds, direct contact, and the discharge of sewage sludge and effluent to land or water environments. Similarities between human and animal MRSA lineages (Cuny et al., 2010) and apparent enrichment or spread of transferable AMR during sewage treatment (Silva, Castillo, Callejas, Lopez, & Olmos, 2006) are examples of interesting phenomena in this area. The epidemiological investigation by Grøntvedt et al. (2016) clearly demonstrated the potential for workers to carry MRSA of the livestock-associated clonal complex (CC398) across national boundaries and to introduce it to pig units.
Control and Elimination of AMR Organisms
The role of vectors
There is good evidence that rodents act as vectors, reservoirs, and possibly multipliers for Salmonella on pig and poultry units and in animal feed preparation facilities (Daniels, Hutchings, & Greig, 2003; Davies & Wales, 2010, 2013; Martelli, Wales, & Davies, 2017; Wales et al., 2009). Wild birds and other wildlife may have a similar, if possibly lesser, role (Andrés et al., 2013; Andres-Barranco et al., 2014; Carlson et al., 2015). Flying insects have been found to carry Salmonella, E. coli O157, and Campylobacter in the vicinity of livestock units, and experimental transmission of the last two agents between animals has been demonstrated using flies as vectors (Wales et al., 2010).
There is also a substantial body of literature demonstrating the carriage of zoonotic bacteria and of AMR in pathogens and commensal bacteria by birds, rodents, and other wildlife, often in proximity to livestock (Greig et al., 2015; Rothenburger, Himsworth, Nemeth, Pearl, & Jardine, 2018). The following examples contain some features of particular interest. Enterococcus faecalis isolates from wild birds in Poland were of diverse subtypes, similar to human and livestock isolates, and were commonly resistant to multiple antimicrobial drugs (St˛ epie´ n-Py´ sniak, Hauschild, Nowaczek, Marek, & Dec, 2017). Among E. coli from Irish deer and herring gulls, AMR was seen in 31% and 87% of isolates, respectively, perhaps to some extent reflecting the relative intensities of interaction with human environments of the two species (Smith, Wang, Fanning, & McMahon, 2014). Gulls have been observed to carry carbapenem-resistant members of the Enterobacteriaceae in several countries, and this is likely to be a result of exposure to human sewage waste (Köck et al., 2018). In Germany a migratory bird (Black Kite) yielded a locally uncommon Salmonella serovar (Corvallis) with plasmid-borne MDR that included resistances against cephalosporins and carbapenems (Fischer, Schmoger, Jahn, Helmuth, & Guerra, 2013). Highly similar subtypes of MRSA have been isolated at high prevalence from both pigs and rodents on pig units (Pletinckx et al., 2013). In this context it is also worth noting that horses have been shown commonly to carry ESBL-expressing E. coli (Sadikalay et al., 2018; Walther et al., 2018), so other livestock or companion animal species may be regarded similarly to wildlife with respect to potential vector status.
The majority of wildlife studies is observational, with few interventional or cohort studies that can properly examine the direction of movement of bacteria between livestock and wildlife (Greig et al., 2015). Among studies where the direction(s) of influence can be inferred, better rodent control (or at least a smaller rodent population) was associated with better control of environmental-persistent Salmonella in hen houses (Martelli et al., 2017), while a substantial reduction of the prevalence of Campylobacter-colonized broiler flocks (50% to 75% compared with baseline or control data) has been observed in the warmer months in Denmark and Iceland by fitting fly screens to poultry houses to prevent ingress of insects (Bahrndorff, Rangstrup-Christensen, Nordentoft, & Hald, 2013; EFSA, 2011a).
Therefore, fair evidence exists that for bacterial genera of concern with respect to AMR, there is a likely role for wildlife and insects acting as mechanical or biological vectors, introducing resistant organisms onto premises and/or contributing to spread or carryover between groups of animals. Furthermore, migratory birds may transfer AMR organisms over considerable distances. Consequently, there are aspects of biosecurity relating to wildlife, both external (for example, excluding wild birds from feed stores) and internal (for example, rodent control), which are likely to assist the control of AMR. Indeed, Taylor et al. (2016) identified wild bird access to poultry houses as a risk factor for FQ resistance among Campylobacter in poultry. However, robust data in this general area are lacking at present.
Fitness effects of AMR
AMR often imposes physiological costs on bacteria, whether by elevated resource demands or interference in biochemical processes, such that a selective pressure may be needed to maintain AMR in a population. Many AMRs will decline following cessation or marked reduction in the use of relevant drugs, such as has been observed among Dutch isolates of E. coli and Campylobacter in the last decade following stringent control of antimicrobial use in agriculture (RIVM, 2017, p. 80). However, by contrast, FQ resistance among isolates of Campylobacter in North America and Denmark has remained prevalent despite little or no recent use of FQ in the respective industries (Agunos et al., 2013; DANMAP, 2017, p. 130)
Many resistance-conferring mutations appear to have little or no fitness cost (Sander et al., 2002), and compensatory mutations may obliterate any costs or indeed confer a fitness advantage (Schrag, Perrot, & Levin, 1997) in some environments (Basra et al., 2018). Some resistances are at least partly mediated by constitutive efflux mechanisms, for which the machinery of the cell is already well-adapted (Maillard et al., 2013). Experimentally, a topoisomerase (parC) mutation in E. coli was associated with both enhancement of preexisting FQ-resistance and a shift from fitness cost to a fitness advantage, suggesting that against a background of reduced susceptibility, selection for fitness alone might in some cases increase antimicrobial drug resistance (MDR) (Marcusson, Frimodt-Møller, & Hughes, 2009). Moreover, a fitness disadvantage observed in a laboratory test may not always be seen under field conditions, where competing antimicrobial-sensitive bacteria will commonly be stressed and growth-limited by prevailing environmental conditions. Under conditions of low fitness costs (or of fitness benefits), not only may antimicrobial-resistant bacteria persist in the absence of selective pressure, but the barrier to the introduction and establishment of such strains on premises from outside may be minimal.
Pertinent examples where little or no fitness cost has been observed experimentally (in vitro and/or in vivo) in resistant organisms include: FQ-resistant Campylobacter (Zhang, Lin, & Pereira, 2003), FQ-resistant human E. coli (de Lastours et al., 2014), colistin-resistant Salmonella (Rhouma et al., 2016), and E. coli after introduction of a MDR plasmid (Enneetal., 2005). Resistance plasmids appear in some cases to carry elements that can compensate for the cost of resistance they encode, as in the case of a colistin-resistance (mcr-1) plasmid in E. coli (Tietgen et al., 2017). Assessment of E. coli strains bearing ESBL-encoding plasmids did not identify a consistent fitness cost to the plasmid host in terms of growth or metabolic activity compared with plasmid-cured variants, and indeed some of the plasmid-bearing wild-type strains had superior biofilming capability, which may be important in environmental survival (Schaufler et al., 2016).
In consequence of the foregoing, although antimicrobial use on farms clearly affords many AMR organisms an advantage, removal of that selective pressure may not be sufficient alone (even over an extended period) to effect a substantial reduction in the prevalence of some AMR. Such effects are likely to vary between organisms and resistances but the removal of resistant organisms by C&D, and the prevention of new resistant organisms being brought in and spreading, are likely to be important tools alongside reduction of antimicrobial use.
Possible role of animal feed
It is recognized that animal feed is at risk of contamination by microorganisms, including enteric bacteria such as E. coli and Salmonella that can carry and transmit AMR. These may enter the feed from source materials, by endemic or periodic contamination at processing facilities, by postprocessing contamination via transport and storage facilities, plus wildlife access and/or by contamination in feeders (Davies & Wales, 2010, 2013). Relevant on-farm biosecurity measures include attention to standards of feed supplies, and to feed unloading and storage facilities (Ghent University, n.d.).
There are a few reported studies that specifically examine AMR in animal feed and ingredients. Feed mill sampling in Portugal yielded viable E. coli in 50% and 30% of poultry feed and ingredient samples, respectively, with Enterococcus spp. grown from approximately twice as many samples (da Costa, Oliveira, Bica, Vaz-Pires, & Bernardo, 2007). AMR, often multiple, was common among both the E. coli and the enterococci. The examination of feed on farms for AMR has found, for example, ESBL- and AmpC-producing E. coli in broiler feeders but not in stored feed (Dierikx et al., 2013), and MRSA in pig feed where the precise location of feed was not specified (Pletinckx et al., 2013). A study in Iceland found 32% of E. coli in samples from broiler feeders to be FQ-resistant (Thorsteinsdottir, Haraldsson, Fridriksdottir, Kristinsson, & Gunnarsson, 2010), but whether the farm samples had potentially been in contact with birds before sampling is unclear.
Therefore, given the known risks of external contamination of feed by certain bacteria and of on-farm feed contamination by wildlife (Davies & Wales, 2013), there is likely to be a risk (albeit unquantified at present) of AMR posed by feed biosecurity breaches. Hygiene practices around feed stores, conveyers, and dispensers on farm are also of potential significance for persistence and carryover of resistant bacteria. Feed residues, particularly when moistened by incompletely effective wet C&D procedures, provide suitable environments for the survival and growth of many bacterial species (Wales, Breslin, & Davies, 2006). Certain animal-related feed ingredients such as feather meal are conceivably a source of selective concentrations of antimicrobial substances even when harvested beyond conventional antimicrobial drug withdrawal times (Chiesa, Nobile, Panseri, & Arioli, 2018). The use of such materials in animal feed is currently heavily restricted by the European Union (Regulation (EU) Nr 56/2013). Apart from livestock feed, the recent marked rise in popularity of feeding pet cats and dogs a raw diet poses another risk of the introduction and dissemination of AMR. On livestock units such animals may be used for stock management, pest control, or as companions. AMR bacteria (E. coli or Salmonella) have readily been isolated from uncooked pet food or treats (Baede et al., 2017; Nilsson, 2015; Pitout et al., 2003). Furthermore, raw feeding has been identified as a risk factor for the fecal shedding of: multidrug resistant E. coli (Groat, Williams, Pinchbeck, Simpson, & Schmidt, 2016, p. 540), E. coli resistant to extended-spectrum cephalosporin antimicrobials (Baede et al., 2017; Groat et al., 2016, p. 540; Lefebvre, Reid-Smith, Boerlin, & Weese, 2008), and antimicrobial-resistant Salmonella (Leonard et al., 2015).
Examples of control schemes in Europe
LA-MRSA in Norway. LA-MRSA was first identified in Norway in 2011, in pigs. Subsequent surveillance identified three clusters of MRSA-positive pig farms in 2013/2014 (Grøntvedt et al., 2016). Farms identified as contaminated have been subjected to depopulation, thorough C&D of premises, sampling of the environment to demonstrate effective removal of the organism and mandatory downtime before restocking with pigs from MRSA-negative herds. This regime, although described as "expensive and time-consuming for all involved" (Norwegian Veterinary Institute, n.d.), has resulted in just nine MRSA-positive farms (not all having the livestock-associated subtype) identified since 2014 in surveillance of between 821 and 986 herds annually, targeting all genetic plus commercial sow herds (Urdahl, Norstrom, Bergsjø, & Grøntvedt, 2018, p. 6). From these, contact tracing has identified a further 63 MRSA-positive units. It is reasonable to say that the national cost of implementing such a program in countries with much larger pig industries than Norway would be daunting, given the proportion of pig units that might need to be depopulated.
Reduced antimicrobial drug use and AMR in Netherlands. Partly in response to the emergence of LA-MRSA in pig herds, plus the identification of prevalent ESBL E. coli in broiler meat, in the last decade Dutch government action has become focused on reducing the use of antimicrobials across the agricultural industry (SDa, 2017, p. 95). The taskforce and Health Council recommendations from 2008 onward have been implemented by: enhancing the monitoring of AMR and antimicrobial use, clarifying the definitions of farmers' and vets' responsibilities and relationships, and requiring individually tailored farm health plans and treatment plans. Latterly, restrictions have been introduced on the use of FQ or of third- and fourth-generation cephalosporins in farm animals, as well as on farmers' authorization to administer antimicrobials autonomously. It has been observed that substantial reductions in antimicrobial drug use only occurred once memoranda and plans were enforced by governmental regulations and targets, but that implementation on farms has remained a matter of individual and private assessments and arrangements (Speksnijder et al., 2015). The effect of the strategy in the Netherlands has been a substantial reduction in agricultural antimicrobial drug use (50% between 2007 and 2012, with continued reductions since), with a much greater reduction in the use of FQ and higher-generation cephalosporins (RIVM, 2017, p. 80; Speksnijder et al., 2015). Reductions achieved have varied substantially between sectors, partly associated with baseline usage (SDa, 2017, p. 95). Correlated with this reduction, AMR has declined among E. coli from broilers, veal calves, and pigs, and in FQ resistance among Campylobacter jejuni isolates from poultry (Dorado-Garc´ ıa et al., 2016; RIVM, 2017, p. 80; Speksnijder et al., 2015).
Reduced antimicrobial drug use in Denmark. Denmark has adopted an alternative approach, mixing regulation with voluntary initiatives, but still employing detailed surveillance of antimicrobial usage and AMR to assist and quantify the impact of interventions (DANMAP, 2017, p. 130; Frandsen & Kornholt, 2012). Danish agricultural antimicrobial use is heavily skewed (80%) toward pig production. Regulatory restriction on FQ achieved an 84% reduction in use between 2001 and 2005, but in the face of increasing overall antibiotic use in the pig industry despite guidelines introduced in 2005, a "yellow card" scheme was introduced in 2010. This penalizes farms that exceed a threshold standardized use of antimicrobials, by restricting further use and mandating veterinary review. The scheme has recently been further refined, weighting usage by class of antimicrobial. In addition, the pig and dairy industries have introduced voluntary bans on the use of cephalosporins.
Overall antimicrobial use in the Danish pig sector, standardized to an equivalent proportion of population treated per day, has reduced by 27% since 2009 (DANMAP, 2017, p. 130). Resistance monitoring of E. coli (150 to 300 abattoir isolates annually) from Danish pigs has not shown any clear trend overall since 2001, although there has been a gradual increase in the last decade in resistance to ampicillin, trimethoprim, and tetracycline. Importantly, resistance to FQ and to third-generation cephalosporins has remained very low (DANMAP, 2017, p. 130).
Reduced antimicrobial drug use in the United Kingdom. Measures to radically reduce antimicrobial use in U.K. agriculture have the weight of public opinion behind them, and progress has been achieved in reducing agricultural use since the government's AMR strategy commenced in 2013 (HM Government, 2017). Currently, the strategy is focused on voluntary action promoted by government and organizations within each industry sector, latterly backed by sector-specific targets (RUMA, 2017, p. 59). Recent overall reductions (in milligrams antimicrobial drug per kilogram standardized biomass) have been substantial: 8% in 2015 over the 2013 baseline, and 21% from 2015 to 2016, with reductions in the latter period of around one-third for the pig and broiler sectors (VMD, 2017, p. 98).
If further controls are deemed necessary, it may be easier to gain general acceptance for (and therefore to introduce) a regime such as that of Denmark, targeting higher antimicrobial-use farms and seeking the complete avoidance of some antimicrobial classes via industry schemes, rather than a Dutch model of intervention across the board, backed by regulatory compulsion. Either approach requires reliable and comprehensive data on antimicrobial use at an individual farm level for effective operation. In the U.K. pig and poultry sectors such data are increasingly available, while elsewhere sufficient detail is currently lacking (VMD, 2017, p. 98).
Disinfection
Efficacy against the main antimicrobial-resistant organisms
Some bacterial taxa, such as mycobacteria or spore-forming organisms, are especially resistant to destruction and elimination from premises by disinfection. However, these do not include the bacterial groups that currently pose livestock AMR challenges: principally staphylococci, streptococci, enterococci, spirochetes, Campylobacter spp., and members of Enterobacteriaceae. Such organisms are assumed to be susceptible to disinfectants in formulations and at concentrations approved in the United Kingdom by the Dept. for Environment, Food and Rural Affairs (Defra) for General Orders use, where the test for efficacy uses Salmonella enteritidis as the reference organism (UK Government, n.d.).
Nonetheless, challenges to effective C&D are not trivial and they include: adequate and comprehensive removal of organic soil; ensuring complete coverage of surfaces for sufficient contact time with disinfectant of appropriate concentration; avoiding interfering substances in diluents or detergents used before disinfection; access to cracks, recesses, and voids in buildings; effects of organic matter on disinfectant action, and bacterial growth in moist environments where disinfection has been subtotal (Davies & Wray, 1995; Stringfellow et al., 2009; Wales et al., 2006). Given the well-documented difficulties in removing bacterial pathogens from animal accommodation (Boughton, Egan, Kelly, Markey, & Leonard, 2007; Carrique-Mas, Marin, Breslin, McLaren, & Davies, 2009; Mannion, Lynch, Egan, & Leonard, 2007; MuellerDoblies, Carrique-Mas, Sayers, & Davies, 2010; Wales et al., 2009), it cannot be assumed that AMR organisms will be any easier to eliminate. Indeed, in a hatchery, on broiler farms and in an abattoir lairage, AMR Salmonella and/or E. coli have been found to persist after C&D (Bolton, Ivory, & McDowell, 2013; Taylor et al., 2016).
It is usually assumed that conventional C&D procedures aimed at eliminating an environmentally robust nonspore-forming pathogen such as Salmonella will be similarly effective against AMR organisms on farms. The challenge has been regarded in both cases as a technical one, of using suitable and effective methods, yet with conventional disinfectant concentrations and contact times. However, there is some evidence (Kotb & Sayed, 2015; UK Animal and Plant Health Agency [APHA] unpublished laboratory trials) that LA-MRSA can be more resistant than Salmonella to elimination by disinfectants and is not controlled by approved concentrations of some commonly used disinfectants, even in bacterial suspension tests. This may prompt consideration of more robust disinfection conditions for this AMR organism at least, perhaps using assessments focused on the particular organism in question. It also may lead to questioning of assumptions made about other AMR organisms, such as ESBL-producing E. coli or Campylobacter with efflux pump and chromosomally mediated resistance to FQs.
Co-selection of AMR by disinfectants
Compared with antimicrobial drugs, disinfectant preparations are not required to be minimally toxic to animal tissues at normal use concentrations, but conversely they do need to operate within comparatively short timescales, often at cool temperatures, and to be microbicidal without assistance from the immune system. Therefore, they usually target more than one element of the bacterial cell. Cellular targets can be quite nonspecific (for example, disrupting cell membrane structure or coagulating cytoplasmic proteins) rather than binding specific enzymes or functional proteins, and typically disinfectants are used at several multiples of their MICs to achieve a rapid kill effect (Wales & Davies, 2015). Despite this focus on an overwhelmingly lethal effect, the conditions of use of disinfectants in the field permit some microorganisms to be exposed to much lower concentrations of the active components. This may occur where there are interfering substances present (for example, residual organic soil, biofilm matrix, or suspended organic material in diluent), when puddles or moisture films preexist on surfaces (for example, from washing procedures), through technical errors or inappropriate economizing with in-use dilutions, or when waste disinfectant or rinsing water is disposed of into drains, slurry stores, or allowed to soak into soil. Under these conditions, bacteria potentially could be selected for decreased susceptibility to certain disinfectants.
Field studies have provided some evidence for reduced susceptibility to certain disinfectants among bacterial isolates from hatcheries (where disinfectants are applied daily ,usually at sublethal concentrations) and biocide manufacturing plants (Lear, Maillard, Dettmar, Goddard, & Russell, 2002; Willinghan, Sander, Thayer, & Wilson, 1996). However, other studies (examining farm bacteria) have failed to find significant evidence of such associations (Aarestrup & Hasman, 2004; Martin & Maris, 1995). Findings with isolates from human medical settings are similarly inconsistent (Wales & Davies, 2015). Experimentally, controlled exposure of laboratory and field strains of bacteria, including members of the Enterobacteriaceae and Staphylococcus spp., to subinhibitory or gradually increasing concentrations of disinfectant can be associated with decreased susceptibility to the biocide in question, although the degree to which this happens and the stability of the acquired change in susceptibility varies substantially from case to case (Bloomfield, 2002; Wales & Davies, 2015). Field and experimental evidence, discussed in Which Disinfectants are Most Problematic in This Respect? section, both point to important differences between classes of disinfectant with respect to the risk of reduced susceptibility among target organisms, and of associated mechanisms.
Whereas a specific, single-step change can result in bacterial resistance to an antimicrobial drug, similar mechanisms are seen only rarely with biocides. The mutation and upregulation of fabI conferring reduced susceptibility to triclosan, a food and personal hygiene product (Rensch, Klein, & Kehrenberg, 2013) is a prominent example, but of limited relevance in the field of livestock disinfection. Such effects are considered likely to have little effect in "real world" disinfection where application concentrations are typically multiples of the lethal dose and several targets are affected simultaneously (Bloomfield, 2002). However, there are some phenomena that confer reduced susceptibility both to biocides and to antimicrobial drugs (co-selection), and which are either part of the normal adaptive repertoire of microorganisms (intrinsic) or are readily acquired by mutation or genetic transfer under appropriate conditions.
Intrinsic resistance may be exhibited following phenotypic adaptations to the environment (such as biofilm formation or nutrient stress responses), or be associated with adaptation to survival in relatively protected environments, such as phagocytes in the body or protozoa in the soil (Bloomfield, 2002; Gilbert & McBain, 2003). By contrast, acquired or adaptive resistance is associated with changes in the organism caused by mutation and/or altered expression of endogenous genes, or by transfer of mobile exogenous genetic material (Russell, 1995; Tumah, 2009). Adaptation may be a constitutional and essentially nonreversible effect, or it may be a transient and regulated response to conditions.
Co-selection between reduced disinfectant and antimicrobial susceptibilities of bacteria may occur because a structural or metabolic change conferring reduced susceptibility to a disinfectant also affects the action of one or more antimicrobial drugs. This is termed cross-resistance and often happens in association with changes to the cell envelope, where there can be changes in permeability and/or in efflux (Buffet-Bataillon, Tattevin, Bonnaure-Mallet, & Jolivet-Gougeon, 2012; Chapman, 2003; Maillard et al., 2013). Alternatively, the linkage may be genetic, where a disinfectant-resistance feature (such as structural or regulatory genes for efflux, or an enhanced biofilm capability) is encoded on the same mobile element as AMR genes, and thus they are transmitted, acquired, and selected for together. This is termed co-resistance (Chapman, 2003; Maillard et al., 2013). A common example relevant for livestock is the co-location of qac genes (conferring reduced susceptibility to QACs) and genes encoding resistance to antimicrobial drugs, in bacterial species including staphylococci and members of the Enterobacteriaceae (Buffet-Bataillon et al., 2012; Jaglic & Cervinkova, 2012).
Global regulatory responses may drive cross-resistance (Karatzas et al., 2008) and these changes may be stress responses (typically reversible) or mutational shifts. A well-studied example of the latter is the multiple antibiotic resistance (MAR) phenomenon in E. coli and other members of the Enterobacteriaceae. This is a common phenotype associated with AMR or controlled exposure to biocides (Russell, 2000). On its own it generates modest increases in MIC values (around four- to eightfold) for a range of antimicrobial drugs (Morente et al., 2013). Sensitivity to organic solvents, triclosan, and pine oil are characteristically decreased, but effects on other biocide MIC values vary according to the particular biocide or biocide mix, and there may indeed be no significant change (Karatzas et al., 2008). The phenomenon arises following mutational or substrate-mediated inactivation of the MarR transcriptional repressor, leading to increased expression of the MarA global regulator and causing, among other things, upregulation of the mar operon itself plus the AcrAB-TolC multidrug efflux pump, and downregulation of the OmpF porin (Grkovic, Brown, & Skurray, 2002; Oethinger, Podglajen, Kern, & Levy, 1998; Putman, van Veen, & Konings, 2000).
A further mechanism that may be relevant is the influence of exposure to sublethal concentrations of disinfectant on rates of genetic transfer. Experiments involving field (sewage) bacterial communities and laboratory strains of E. coli and S. aureus show that the efficiencies of conjugation and transduction may be enhanced, unaffected, or reduced in the presence of disinfectants, depending on the agent used and its concentration (Jutkina et al., 2018; Pearce, Messager, & Maillard, 1999; Zhang, Gu, He, Li, & Chen, 2017). Furthermore, given that low concentrations of antimicrobial drugs elevate the rate of random mutations in exposed bacterial populations (Cogliani, Goossens, & Greko, 2011), it is reasonable to suppose that a similar phenomenon may occur with disinfectants, which may consequently increase the generation of (as well as selection for) spontaneous mutants showing cross-resistance to disinfectants and antimicrobial drugs.
Selection of antimicrobial-resistant and biofilm-forming organisms or spontaneous resistant mutants from mixed populations exposed to disinfectants
Because of the phenomenon of co-selection of disinfectant and AMR, disinfectant use on livestock premises and equipment potentially can support or select for some AMR bacteria. The selective pressure of sublethal disinfectant exposure may nullify a fitness disadvantage or indeed confer a comparative advantage in such organisms. As an example, a bacterial cell that shows highlevel or multiple antimicrobial drug resistance may have reduced membrane permeability and mutational upregulation of a multidrug efflux pump. These features can reduce the cell's access to nutrients and increase its energy requirements, thus limiting its growth potential, but they could also confer relative protection in the face of disinfectant exposure. Alternatively, if there is little or no fitness cost to the AMR, but the genetic element conferring it is linked to, for example, an adaptive efflux or biofilm capability, then disinfectant selective pressure could actually increase the relative proportion of AMR organisms, rather than just promote their survival.
In field conditions, biofilms may have a major role to play in bacterial survival of C&D procedures. One study showed hatchery environmental isolates of E. coli to have superior biofilm characteristics compared with contemporaneous meconium isolates (Osman et al., 2018). Biofilms are comprised of an extracellular matrix that provides a diffusion barrier and an enhanced medium for bacterial signaling and genetic exchange, in addition to being a potential site for neutralization or binding of disinfectants: enzymes degrading triclosan, chlorhexidine, and some oxidizing agents, among others, are known to occur within biofilms (Bridier, Briandet, Thomas, & Dubois-Brissonnet, 2011; Gilbert, Allison, & McBain, 2002; Maillard et al., 2013; Morente et al., 2013; Tumah, 2009). Minor elevations in MIC may have increased significance for survival of a bacterium in the context of a biofilm diffusion gradient (McBain & Gilbert, 2001). Furthermore, bacteria in biofilms show increased mutability and are relatively nutrient-deprived, exhibiting stress responses that additionally diminish their susceptibility to chemical attack (Bridier et al., 2011; Gilbert et al., 2002; Maillard et al., 2013; Russell, 2003).
There is evidence for protection of both gram-positive and gram-negative bacteria by biofilm, and elevated MIC values in the order of 10- to 1000-fold have been observed for a QAC (benzalkonium chloride) and triclosan, among other biocides, with respect to activity against biofilmed versus planktonic veterinary-relevant bacteria, such as E. coli, Salmonella spp., and S. aureus (Morente et al., 2013; Tumah, 2009). In the field, biofilms are heterogeneous bacterial communities (with characteristics distinct from laboratory-grown monoculture films), therefore enhanced and cross-species disinfectant protection may occur (Bridier et al., 2011). In one example, thick biofilm formed by a persistent hospital Bacillus subtilis strain protected both itself and S. aureus from peracetic acid disinfectant (Bridier et al., 2012).
ESBL-bearing plasmids in E. coli have been observed in association with altered biofilm capability (Schaufler et al., 2016). This potentially is an example of antimicrobial/disinfectant coresistance, although disinfectant susceptibility under field conditions has not been reported in these strains. Consequently, biofilm capability may influence the environmental persistence of ESBL/AmpC-bearing and FQ-resistant E. coli, as has been documented in hatcheries (Dierikx et al., 2013).
Investigations of bacterial strains recovered from the field have yielded some evidence of associations between biocide use and reduced susceptibilities to antimicrobial agents. Elevated MIC values for QACs were frequently observed among E. coli recovered from meat in the United States, where such agents commonly are used as sanitizers (Zou et al., 2014). A strong association was found between the frequent disinfection of pens and colonization of nursery piglets with MRSA, with isolates universally encoding genes (qac, smr) responsible for reduced susceptibility to QACs (Slifierz, Friendship, &
Weese, 2015).
However, it is important to note that, despite the mechanisms identified above that may promote the survival and selection of AMR by disinfection, there is as yet no compelling field evidence that in general antimicrobial-resistant bacteria hold a survival advantage during C&D (Oggioni, Furi, Coelho, Maillard, & Martinez, 2013; Wales & Davies, 2015). This may reflect the fact that experimentally reduced disinfectant susceptibility in bacteria is often unstable and is associated with highly variable effects on AMR, including, in some cases, reversion to susceptibility (Curiao et al., 2015; Wales & Davies, 2015). The selection of spontaneous antimicrobial-resistant mutants by disinfectants in the field has not yet been addressed experimentally.
Which disinfectants are most problematic in this respect?
As already discussed, disinfectants typically have nonspecific effects and are applied at concentrations which are aimed to achieve a rapid kill. However, the bisphenol agent triclosan relies substantially on interaction with a precise target (enoyl acyl carrier protein reductase; fabI and homologues) at normal working concentrations (Carey & McNamara, 2015). The biocidal effect is thus susceptible to interference by mutation plus upregulation of fabI expression (Rensch et al., 2013) and, moreover, the kill effect diminishes rapidly with declining effective concentration (Maillard et al., 2013). Reduced susceptibility to triclosan in bacteria exposed to sub-MIC concentrations of the agent tends to be substantial and stable, in contrast to many other disinfectants including chlorhexidine and hydrogen peroxide (Wesgate, Grasha, & Maillard, 2016). However, a key feature of such investigations is a high degree of variation between differing combinations of agent and bacterial strain or species (Braoudaki & Hilton, 2004; Wales & Davies, 2015).
QACs are a widely used group of biocidal agents, many of which are used as disinfectants. They disrupt the structure and function of bacterial membranes, and their popularity reflects their versatility and safety profile. However, it is known that use of a QAC can select for QAC-resistant bacteria, mediated by efflux pumps (plasmid-encoded or upregulated in response to exposure) and/or membrane changes, in addition to promoting biofilm formation (Buffet-Bataillon et al., 2012; Buffet-Bataillon, Tattevin, Maillard, Bonnaure-Mallet, & Jolivet-Gougeon, 2016; Tansirichaiya et al., 2017; Wales & Davies, 2015). Furthermore, allied reductions in antimicrobial sensitivity have been observed. Notably, these include FQ resistance associated with efflux, mutations in the chromosomal Quinolone Resistance Determining region, and increased genetic exchange (Buffet-Bataillon et al., 2016). In this context, there are concerns about the increasing use of QAC (benzalkonium chloride) disinfectants in poultry hatcheries, at sublethal concentrations as a consequence of cost and staff safety issues. Certain quinolone-resistant hatchery-resident Salmonella strains appear to have emerged from such situations and subsequently spread to broilers (APHA unpublished data: Salmonella in Livestock Production in GB, 2018 report).
An allied concern of particular relevance to farms is the relative environmental persistence of many disinfectants such as QACs and triclosan, and their tendency to bind to organic matter and soil (Buffet-Bataillon et al., 2012; Hegstad et al., 2010). This provides a potential long-lasting low-level exposure of microbes to selective pressure. Concentrations of triclosan in biosolids and sediments have been reported to lie between the MIC of "fully susceptible" and "tolerant" bacteria (Carey & McNamara, 2015). Sampling of watercourse sediment downstream of a plant discharging QACs showed an increased proportion of environmental bacteria to be harboring QAC efflux genes (qacE), in association with class 1 integrons, plus a shift toward gram-negative bacteria (Gaze, Abdouslam, Hawkey, & Wellington, 2005). Class 1 integrons are associated with carriage and horizontal transfer of AMR gene cassettes.
Where QAC disinfectants fail to eliminate bacteria that have developed a reduced susceptibility, alternative products are needed. In many circumstances oxidative preparations based on potassium peroxymonosulfate appear to be suitable and effective. Such compounds are infrequently associated experimentally with stable decreases in bacterial susceptibility to disinfectants or to antimicrobial drugs (Wales & Davies, 2015), and field experience bears this out. As a class, they do potentially suffer from reduced effectiveness in the presence of biofilm and organic soil, owing to quenching by extracellular components (Chapman, 2003; McLaren, Wales, Breslin, & Davies, 2011), but this can be combated by improved cleaning in cases where this is considered to be an issue. Unfortunately the use of these agents in hatcheries is limited because their corrosive effects compromise both staff safety and the expensive equipment used for chick incubation and processing (Samberg & Meroz, 1995).
C&D properly to eliminate antimicrobial-resistant organisms
The elimination of bacteria harboring AMR from premises, transport vehicles, and equipment such as poultry crates is a challenge similar to the elimination of pathogens such as Salmonella. Factors that in general are controllable include: selection of a suitable disinfectant, dilution to a correct concentration with clean water, and achieving a specified contact time. Thoroughness of cleaning is also substantially a consequence of the time and care taken (and the frequency with which cleaning is performed), but surface deterioration, building design, and the presence of immovable equipment such as feeders and cages can make some residual soiling almost inevitable in certain cases. Other factors such as ambient temperature are also difficult to control.
It has been recognized on farms that wet cleaning (washing down) can sometimes be associated with poorer results in terms of pathogen or AMR control than dry-cleaning using scrapers, brushing and vacuum removal of dirt and dust (Taylor et al., 2016; Walesetal., 2006). This is likely to reflect spreading by aerosols and growth of residual bacteria after exposure to moisture, especially if embedded within organic media, and possibly some diluting effect on subsequently applied disinfectant agents.
It is probably unrealistic to expect a single cycle of C&D to reliably eliminate even nonsporulating bacteria from a complex environment such as livestock premises, particularly as intestinal commensal organisms are typically present in much higher numbers than pathogens such as Salmonella. The 5 log-unit reduction that is required to pass Defra-approval tests, even if consistently achievable in field situations (which is doubtful), would leave some residual Enterobacteriaceae, including AMR organisms. However experience, for example, in Sweden with Salmonella in pigs, has shown that if introduction of new strains are prevented and infected stock are identified and removed, intensive C&D can over time lead to the elimination of Salmonella from premises (Osterberg, 2010). Similarly, depopulation and effective C&D methodology has successfully removed LA-MRSA from Norwegian pig units (Grøntvedt et al., 2016).
Disinfection agents vary in their capacity to eliminate or markedly reduce bacterial counts in cleaned livestock accommodations (McLaren et al., 2011; Wales et al., 2006), but the choice of a suitable agent may be governed by cost, safety, and licensing considerations, plus practical issues such as the possibility to effectively seal buildings for fumigation or fogging procedures. Follow-up bacteriological sampling, particularly of locations such as feeders, visibly soiled areas, and defects in surfaces, can verify effectiveness and help to direct future efforts at elimination.
Besides C&D, farms may have difficulties in the suitable disposal of waste, including wash water created as part of the C&D process. Antimicrobials in feces, water, urine, and milk can potentially select for AMR in the environment. Calves fed on milk from antimicrobial drug-treated cows had a higher risk of carrying FQ-resistant E. coli (Duse et al., 2015), and on U.K. dairy farms with ESBL E. coli isolated from cattle, sampling of drains, muck heaps, and slurry recently spread on fields commonly identified ESBL-positive E. coli (Snow et al., 2012). Furthermore, given the tendency of disinfectants such as QACs to persist in soil, the appropriate disposal of unused disinfectant or washing-down water has an AMR-risk aspect to it also. In pig production, the supplementation of feed with copper and/or zinc has led to concern over co-selection of AMR, and livestock farming is considered to be a major source of environmental contamination by these heavy metals (Nicholson, Smith, Alloway, Carlton-Smith, & Chambers, 2003). Unlike disinfectant molecules, these biologically active heavy metals do not biodegrade over time. In consequence of these and other concerns, within the European Union the medicinal use of zinc oxide in feed is being phased out (Anon., 2017) and maximum inclusion levels of copper are being reduced (European Commission, 2018).
Summary and Conclusions
In the field of livestock AMR it is often assumed that there is a relationship betweenbiosecurity and AMR on farms, but there are only a few formal published studies that have attempted to identify such a relationship and to provide evidence of the most important elements. By and large, the studies that have been performed (on E. coli in poultry, pigs, and cattle; on Campylobacter in poultry and pigs; and LA-MRSA in pigs) have supported the experimental hypothesis that the risk of AMR occurring and/or persisting on farms is increased by poor biosecurity, and by the related phenomena of poor hygiene and endemic disease. A correlation has also been noted in some (but not all) cases between better biosecurity and lower antimicrobial drug usage, although antimicrobial usage is undoubtedly an area subject to other major influences. Where technical and economic performance has been assessed in relation to biosecurity and antimicrobial usage (in pig production), the evidence does not indicate that higher production is associated with higher antimicrobial use. However, the evidence base in all these matters is small.
Detailed investigations have been performed into some particular areas of AMR concern, which can provide a narrative view of likely relationships. With respect to AMR among broiler-associated E. coli in vertically integrated Western European production, the roles of imported elite breeding stock and of hatcheries are highlighted in several studies for the introduction and maintenance/transmission of resistant bacteria, respectively. Among young cattle, ESBL multidrug resistant E. coli appear able to colonize readily, and also in the absence of selective pressure, although this may be largely transient without the influence of antimicrobial drugs. In pigs, LA-MRSA appears to travel between layers of the breeding pyramid, but also readily engages in local intra- and inter-species spread. In all these cases, external biosecurity and hygiene (including C&D) are of major importance for control.
There is plentiful evidence that antimicrobial-resistant bacteria are carried by other farm animals (such as horses), by farm wildlife, and by more wide-ranging free species, and that birds in particular may provide a link for resistant organisms between centers of human and livestock habitation. This should prompt biosecurity measures to limit or prevent contact between wildlife and livestock, but the magnitude and direction(s) of wildlife-associated transfers are poorly understood, as is the transfer of AMR from human sources to livestock. Similarly, evidence has emerged of AMR in animal feedstuffs at feed mills and on farms. Thus far, integrated and prospective studies examining possible transmission to animals receiving processed feed, or the effect of countermeasures, are lacking.
There is also a substantial body of experimental data on AMR phenomena such as effects on bacterial fitness, induction of biocide resistance, co-resistance to antimicrobial drugs and biocides, and horizontal transmission of resistance in the presence of antimicrobial agents. Again, very little is known about the significance of these effects and the balance of outcomes in the field, that is, on livestock-associated premises. Assessments of fitness that involve bioassays (Yokobori et al., 2018) or simulations of field environments may be helpful in this area.
The elimination of any environmental bacterial population on livestock premises is challenging. There is, in addition, a very large data gap regarding the effective disinfection of bacteria exhibiting AMR, whether commensal or potential opportunist zoonotic pathogens. An assumption that antimicrobial-resistant bacteria are similar to their nonresistant counterparts in the face of C&D procedures may not be justified, and residual antimicrobial-resistant organisms may enjoy a selective advantage. There is some experimental evidence that this may be the case for LA-MRSA, but there are only few or no relevant data for other organisms. Some environments and C&D regimes (such as in hatcheries) appear wellsuited to the establishment of persistent antimicrobial-resistant populations of bacteria, possibly with enhanced biocide resistance characteristics. Deficiencies in our understanding of biofilms in the field, and in our ability to model them, are of major relevance in this area (Lamas et al.,
2018).
In conclusion, there are important messages emerging from recent data on AMR in livestock. Its development and persistence can no longer be regarded as an issue to be addressed solely by reducing or preventing the use of antimicrobial drugs in agriculture. Biosecurity and hygiene, among other factors, appear to have important roles. However, beyond the prudent and rational use of antimicrobial drugs, data on AMR control are either sparse or largely absent.
For an area of such contemporary significance, there are very few formal, prospective risk factor and longitudinal analytical studies on the occurrence, transmission, and persistence of AMR in livestock and on associated premises. Those studies that do exist point firmly to a role for some aspects of biosecurity, reflecting dissemination pathways that resemble those for zoonotic pathogens such as Salmonella and Campylobacter.
From the evidence we do have there are some important findings that should enhance practical advice on controlling the selection, spread, and persistence of AMR. These include risk factor and tracing data on the spread within and between groups, from treated to untreated animals, and between premises via animals, workers, and vehicles. Consequently, rigorous biosecurity practices should have an impact on AMR. Depending on species and system, these may include: maintaining a closed herd, quarantining incoming animals, screening high genetic value breeding stock for AMR, separation of epidemiological groups plus isolation of sick animals on premises (physically, and by work pattern), and strict controls on personnel access, workwear, and hygiene. C&D to a high standard is likely to provide some control, at least for all-in-all-out systems. Fortunately, many of these measures have also been associated with lower antimicrobial drug use (again, with a modest evidence base), which would also be expected to reduce AMR.
However, there are no meaningful data on the efficacy or selective effect of currently used farm and hatchery products such as detergents, sanitizers, and disinfectants on the survival or emergence of organisms with reduced biocide susceptibility or AMR. This data gap presents difficulties in provision of hygiene advice and may result in the persistence of certain AMR bacteria, despite the removal of selective pressures exerted by antimicrobial medication. Other evidence gaps worth exploring include: the nature and behavior of biofilms in the field regarding C&D, effects of field disinfectants on plasmid transfer, AMR in animal feed, the direction(s) of travel of antimicrobial-resistant bacteria between livestock and wildlife, and the effects of wastes (wash water, slurry, and so on) containing antimicrobial compounds on AMR in the environment. Data from some of these would potentially have wider disease-control applications, beyond AMR in the bacterial species of current concern.
For more of the article, please click here.
Article made possible through the contribution of A. Wales et al.